

【话险危夷】急性呼吸窘迫综合征中肺和膈肌的异质性损伤



目的:在机械通气过程中经常观察到患者与呼吸机不同步,这与较高的死亡率有关。我们假设患者与呼吸机不同步会导致肺和膈肌损伤及功能障碍。
设计:前瞻性随机动物研究
受试者:18只新西兰白兔
干预:通过消耗表面活性物质建立急性呼吸窘迫综合征(ARDS)动物模型;通过膈神经刺激模拟实现分组(辅助控制组、呼吸叠加组和反向触发组)。测量各组肺功能、肺损伤(湿肺与干肺重量比、总蛋白和支气管肺泡灌洗中的白细胞介素-6)、膈肌功能(膈肌力产生曲线)和膈肌损伤(膈肌纤维横截面积、组织学)的影响。呼吸叠加组和辅助对照组(n=2)进行膈肌RNA测序。
方法和主要结果:刺激膈神经以产生较小的吸气力(食道压力波动≈-2.5cmH2O),各组间差异不大。呼吸叠加组的潮气量(>10毫升/公斤)和吸气跨肺压更高,这导致更差的氧合及更差的肺顺应性从而引起肺损伤。反向触发组未引起肺损伤。在辅助控制通气组中没有观察到不同步事件的发生。在呼吸叠加组和反向触发组中出现膈肌偏心性收缩,其中在呼吸叠加组中更常发生。呼吸叠加组和反向触发组膈肌力的产生显著减少。隔肌组织学检查显示,呼吸叠加组异常肌肉的面积比辅助对照组高2.5倍,反向触发组异常肌肉的面积比辅助对照组高2.1倍。膈肌RNA测序分析显示,呼吸叠加组与辅助对照组相比,与肌肉分化和收缩相关的基因受到抑制,而细胞因子和趋化因子介导的促炎反应被激活。
一、前言
自主呼吸是通过呼吸肌收缩进行的自然生理过程,其带来好处包括充分的气体交换和避免了膈肌的萎缩,因此自主呼吸在ICU的机械通气过程中得到了有利促进。随着自主呼吸成为通气管理的核心,医生们发现自主呼吸在急性呼吸窘迫综合征(ARDS)中也可能造成肺部损伤(称为高呼吸驱动肺损伤),其作用机制如肺应激和应变增加、坠积现象和血流灌注量增加。在ICU的机械通气过程中,当自主呼吸增强时,患者呼吸机不同步是常见的,并被认为是高呼吸驱动肺损伤的机制之一。临床观察性研究表明,患者呼吸机不同步与不良结果有关,如较高的ICU、住院死亡率和机械通气持续时间更长,这就产生了患者呼吸机不同步可能导致肺和膈肌损伤,从而导致临床结果恶化的假设。尽管有这一假设看似合理,但没有直接的因果证据支持患者呼吸机不同步对肺部和膈肌造成有害影响。因此,我们建立ARDS实验模型来验证患者呼吸机不同步损伤肺和膈肌的假说。我们模拟了肺损伤兔呼吸机不同步的两种类型,即双侧膈神经刺激引起的“呼吸叠加”和“反向触发”,并用双侧膈神经刺激辅助呼吸,建立“辅助对照”作为对照组。
二、材料与方法
动物准备
将18只新西兰白兔(成年,雄性;3.6±0.1 kg)麻醉并气管造口。插入食道和胃气囊测量食道压力(Pes)和胃压力。
膈神经刺激
识别并暴露双侧膈神经。三组的双侧膈神经刺激是相同的,频率为1.33 Hz(即频率率为80/min),持续0.3秒,并在-2和-3 cm H2O之间获得Pes负偏转的最小可能电压。
实验方案
通过反复灌洗肺直到Pao2/Fio2<150 mm Hg来诱导肺损伤。然后将动物随机分配到三组中的一组(每组n=6)
l辅助控制组;
l呼吸叠加组;
l反向触发组。
对所有动物进行深度镇静,以防止膈肌的自发活动(即除非刺激膈神经,否则Pes不会出现负偏转)。膈神经刺激激活膈肌收缩,诱导每种通气类型。
辅助对照组
动物在容量控制通气(VCV)模式下通气,VT为6-8 mL/kg,频率为40/min(远低于刺激频率),吸气时间为0.35秒,流量触发,呼气末正压(PEEP)为2cm H2O。因此,由双侧膈神经刺激引起的膈肌收缩触发了以80/min的呼吸速率持续4小时的机械呼吸(图1)。该组被设定为对照组。
呼吸叠加组
呼吸叠加被定义为两次短时间的连续呼吸,代表双侧膈神经刺激产生的单一吸气努力。动物采用VCV模式通气,VT为6-8 mL/kg,频率为60/min,吸气时间为0.1秒(远低于刺激持续时间),最小流量触发,PEEP为2cm H2O。由于膈神经刺激(0.3s)超过了预设的吸气时间(0.1s),膈肌收缩触发了两次连续的机械呼吸,呼气时间极短,呼吸叠加模拟了4小时(图1)。
反向触发组
反向触发被定义为在呼吸机启动呼吸之后以及在没有动物启动的辅助呼吸的吸气阶段发生的吸气自发努力。动物采用VCV模式通气,VT为6-8 mL/kg,频率为95/min,吸气时间为0.30秒,PEEP为2 cm H2O。为了避免呼吸叠加随后发生反向触发,流量触发器被调整为不太敏感。由于预设的机械呼吸(95/min)超过了膈神经刺激率(80/min),因此在呼吸机启动呼吸后的吸气阶段,膈肌收缩是随机发生的。这种随机相互作用导致了具有不同起始和终止时间点的各种反向触发类型.
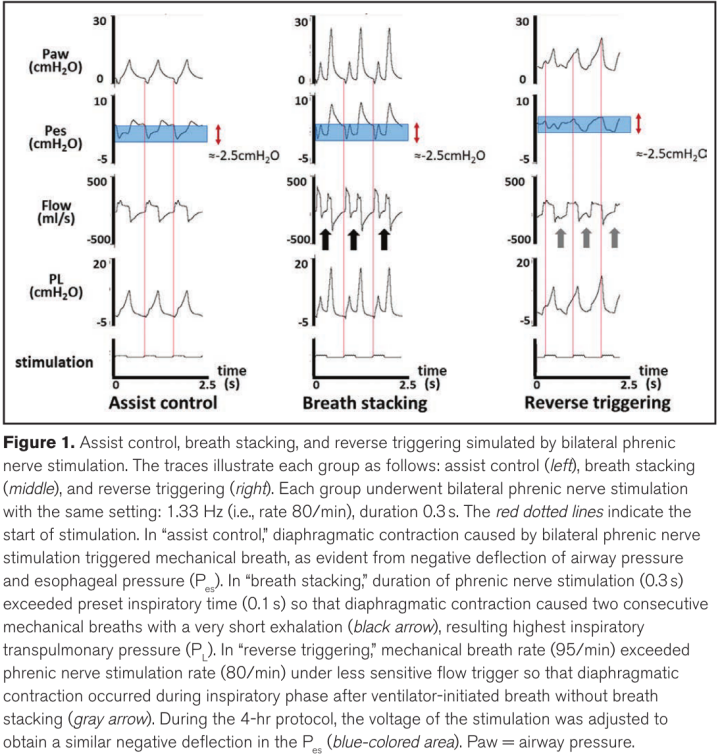
图1。通过双侧膈神经刺激模拟辅助控制、呼吸叠加和反向触发。轨迹显示每组如下:辅助控制(左)、呼吸叠加(中)和反向触发(右)。每组接受相同设置的双侧膈神经刺激:1.33 Hz(即速率80/min),持续时间0.3 s。红色虚线表示刺激开始。在“辅助控制”中,双侧膈神经刺激引起的膈肌收缩触发了机械呼吸,这从气道压力和食道压力(Pes)的负偏转中可以明显看出。在“呼吸叠加”中,膈神经刺激的持续时间(0.3 s)超过了预设的吸气时间(0.1 s),因此膈收缩导致两次连续的机械呼吸,呼气时间很短(黑色箭头),从而产生最高的吸气经肺压力(PL)。在“反向触发”中,超过机械呼吸速率(95/min)
不同步事件
不同步事件使用不同步指数进行量化,定义为不同步事件的数量除以呼吸频率。不同步指数是通过对每个时间点记录的10分钟的值取平均值来计算的。
膈肌力-频率曲线
在基线(肺损伤诱导前)和方案结束时,评估了力-频率曲线,即在每个频率下的跨膈压(Pdi)测量。在10、30、50、80和100Hz的频率刺激双侧膈神经期间,在呼气末对阻塞的气道测量Pdi。所有测量在具有2cm H2O的PEEP进行。
肺部炎症和损伤评估
右肺上叶用于确定湿肺与干肺的重量比。对左肺进行灌洗以测量总蛋白和白细胞介素-6(IL-6)。右肺下叶用于组织病理学分析。
膈肌损伤评估
快速冷冻腹侧膈肌标本,并用苏木精和伊红(HE)和烟酰胺腺嘌呤二核苷酸脱氢酶四氮唑还原酶(NADH-TR)染色。横截面HE染色用于评估膈肌损伤评分。用NADH-TR染色的横截面用于将肌肉纤维分类为I型或II型。
膈肌RNA提取
RNA测序由Macrogen(日本东京)进行。在呼吸叠加和反向触发中观察到相同的不同步事件,即偏心收缩,但频率不同(呼吸叠加中更多,反向触发中更少)。因此,对呼吸叠加与辅助对照进行RNA测序,以研究患有偏心收缩的膈肌转录变化的生物学意义。选择异常肌肉面积分数显示每组平均值的两个样本作为代表。表达谱表示为读取计数和基于每百万个映射读取的外显子的每千碱基片段的归一化值。K-means分类使用iDEP在线软件
统计分析
使用标准软件(SPSS 24)进行统计分析。结果用平均值±标准差表示。重复测量的双向方差分析(ANOVA)用于评估时间和组的影响。重复测量的单向方差分析用于比较力-频率曲线和呼吸参数。所有测试都重复两次,当p值小于0.05时,差异被认为是显著的。
三、结果
呼吸变量
氧合指数(Pao2/Fio2)在“辅助控制”和“反向触发”中随着时间的推移而改善,并且在2小时后大于“呼吸叠加”。在整个方案中,“辅助控制”和“反向触发”具有相似的VT(6-8毫升/公斤),但“呼吸叠加”具有更大的VT(>10毫升/公斤)和更高的呼吸频率(4小时方案),导致1小时和2小时的PaCO2值较低,pH值较高。“呼吸叠加”在整个方案中观察到峰值PL和峰值∆PL最高,从而导致动态呼吸力学和动态肺顺应性在方案实施2小时后最差(附录5,表1)。ΔPes即双侧膈神经刺激产生的自主吸气力,在整个方案中,所有组都相似(≈2.5 cm H2O)。
膈肌收缩力
在“呼吸叠加”中,在方案结束时,与“辅助对照”相比除10Hz(图2)外的所有刺激频率的ΔPdi显著降低(图2);相对于“基线”在除30 Hz以外的其他所有刺激频率上,ΔPdi也显著降低(s图2)。在“反向触发”中,方案结束时,与“辅助控制”相比ΔPdi在所有刺激频率下显著降低(图2),相对于“基线”在除50、80 Hz外的所有刺激频率上显著降低(s-图2)

图2:膈肌力-频率曲线。在方案结束时,将膈肌力的产生评估为每个刺激频率下经膈压力的变化。与“辅助控制”相比,“呼吸叠加”在除10 Hz以外的所有频率下产生的膈肌力更少。“反向触发”在所有频率下生成的膈肌力更少。*p<0.05 vs all,+p值小于0.05 vs“反向触发”
肺损伤
肺损伤在“呼吸叠加”中最大,表现为湿肺与干肺重量比(s-图1A,)、蛋白质浓度(s-图1B)和支气管肺泡液中的IL-6浓度(s-图1C)。“呼吸叠加”观察到支气管肺泡液中湿肺与干肺重量比最高(6.4±1.3;p<0.01 vs所有),总蛋白浓度最高(493±338 mg/dL;p<0.01 vs“辅助控制”,p<0.05 vs“反向触发”),IL-6浓度最高(2.9±2.4 pg/mL;p<0.05 vs“辅助对照”)。各组的组织学肺损伤如图3A所示。
膈肌损伤
各组的组织学膈肌损伤如图示所示(图3B)。直方图显示了每组膈肌纤维(总I型和II型肌纤维、I型肌纤维和II型肌肉纤维)的横截面积分布(见图2)。全面的膈肌纤维在“呼吸叠加”中最大:总肌纤维(4446±684μm2)比“辅助控制”(2636±625μm2;p<0.01)大约70%,比“反向触发”(3415±516μm2;p<0.05)大约30%(2359±538μm2;p<0.01);“呼吸叠加”组Ⅱ型肌纤维(5275±775μm2)大于“辅助控制”组(2925±757μm2;p<0.01)和“反向触发”组(4006±563μm2,p<0.01)

图3。肺和横膈膜的代表性图像。显示了“辅助控制”(左)、“呼吸叠加”(中)和“反向触发”(右)中的代表性图像(原始放大倍数×200)(A:依赖性肺、苏木精和伊红[HE];B:膈肌,HE;C:膈肌、烟酰胺腺嘌呤二核苷酸脱氢酶四氮唑还原酶[NADH-TR])。A、“呼吸堆积”有严重的肺泡损伤,伴有透明膜形成和中性粒细胞浸润。B、在“呼吸叠加”和“反向触发”中,膈肌纤维都受到损伤,其特征是炎症细胞坏死和涌入。C、T型1肌纤维呈现深色(*),而II型肌纤维呈现浅色(+)。“呼吸叠加”同时增大了I型和II型肌纤维,而反向触发仅增大了II型肌纤。
使用点计数技术对三组患者的正常肌肉、异常肌肉和腹膈结缔组织进行比较(图4)。在“呼吸叠加”中,异常肌肉的面积分数(65%±10%)比“辅助控制”高出2.5倍(26%±8%;p<0.01),结缔组织的面积分数最高(vs所有;p<0.01),在“反向触发”中,异常肌肉的面积分数(53%±19%)比“辅助控制”高出2.1倍(26%±8%;p<0.01),而正常肌肉的面积百分比较低(与“辅助控制);p<0.01)。
对于膈肌的RNA测序,通过sd确定的前2000个可变基因进行K-means聚类,并分为三个聚类:聚类A(n=406)、聚类B(n=377)和聚类C(n=1217)(s-图3)。聚类A和B显示了一组基因,其表达在呼吸叠加过程中分别下调和上调。聚类A和B中的GO分析发现,与“横纹肌细胞分化的正调控”、“肌肉收缩”和“肌肉器官发育”的生物学过程相关的基因在呼吸叠加中下调(图5A),而与“细胞因子介导的信号通路”、“对未折叠蛋白的反应”和“对细胞因子刺激的细胞反应”的生物学过程相关的基因在呼吸叠加中上调(图第5B段)。在呼吸堆叠中也观察到与“细胞因子活性”和“趋化因子活性”的分子功能相关的基因的上调(图第5B段)。

图4。腹侧横膈膜的正常肌肉、异常肌肉和结缔组织的比较。正常肌肉、异常肌肉和结缔组织的分布如“辅助控制”(左)、“呼吸叠加”(中)和“反向触发”(右)所示。条形图显示了所有组肌肉损伤的定量得分。“辅助控制”的正常肌纤维数量最高(≈70%)。与“辅助控制”相比,“呼吸堆积”的异常肌肉面积分数明显更高,结缔组织面积分数最高,导致正常肌肉面积分数最低。与“辅助控制”相比,“反向触发”的异常肌肉面积分数显著更高,正常肌肉面积分数明显更低。*p值小于0.05与“辅助对照”,+p值小于0.05vs“全部”。

图5。隔膜的RNA测序。对辅助对照组(AC)和呼气堆叠组(BS)的膈肌进行RNA测序分析(各n=2)。A、 聚类A中的蓝条显示了在BS组与AC组中表达下调的基因组。B、 簇B中的红条显示了在BS组与AC组中表达上调的基因组。分别使用聚类A和聚类B的基因进行生物过程和分子功能项的基因本体论(GO)富集分析。GO生物过程和GO分子功能项按p值排序,并列出前10项。条形图显示–log10(p值)。CCR=CC趋化因子受体。
四、讨论
目前的实验数据表明,密切监测和管理患者与呼吸机的不同步,可能对最大限度地减少ARDS患者自主呼吸对肺和膈肌的损伤很重要。这是因为呼吸叠加会使VT和吸气PL加倍,从而损害肺部,而呼吸叠加和反向触发(无呼吸叠加)会导致膈肌损伤和功能障碍,可能是因为膈肌偏心收缩。
不同步性和肺损伤
这项研究表明,呼吸叠加损伤了肺部,表现为氧合较差、肺顺应性较差、支气管肺泡液中总蛋白和IL-6浓度较高以及肺湿干重比较高。肺部组织学分析证实了这些发现(图3)。呼吸叠加增加肺损伤的一个关键机制是肺泡过度扩张,因为VT更大(≈比目标大1.5×1.5)和吸气PL更高,增加了呼吸机诱导的肺损伤的风险。因此,医生需要意识到呼吸叠加发生频率更高的情况,如呼吸驱动力更高、严重的肺损伤、肺保护性通气期间VT更受限以及吸气时间更短。
我们的数据显示,反向触发不会增加肺损伤(与“呼吸叠加”或“辅助控制”相比)。考虑到既往的观察结果,这一发现似乎出乎意料。对此已经提出了几种解释。
首先,整体VT和吸气PL没有随着“反向触发”(与“辅助控制”相比)而增加。这是因为根据方案,通过减少流量灵敏度的变化,避免了反向触发后的呼吸叠加,并且VCV在反向触发期间保证了与预设值相同的整体VT。值得注意的是,当反向触发发生在伴随叠加呼吸的压力控制或压力调节的容量控制模式期间时,会增加整体VT。在这种情况下,反向触发可能对肺部造成潜在伤害。其次,我们之前的研究表明,它发生在伴随叠加呼吸的压力控制或压力调节容量控制模式下。在这种情况下,反向触发可能对肺部造成潜在伤害。其次,我们之前的研究表明,在反向触发过程中,肺拉伸与膈肌收缩的强度成比例。在本研究中,双侧膈神经刺激引起的微弱膈肌收缩,即≈-2.5 cmH2O的Pes负偏转,可能不足以导致依赖性肺区过度拉伸。反向触发导致支气管肺泡液中总蛋白浓度较高(但不显著)(1.8 倍vs“辅助对照”)和IL-6浓度较高(1.5 倍vs“辅助控制”)。因此,反向触发(无论是否有害)对肺部的影响可能取决于反向触发时整体VT增加的程度和/或反向触发期间横膈膜收缩的强度。
不同步性和膈肌损伤
这项研究表明,呼吸叠加和反向触发都会损伤膈肌,从而导致膈肌无力(图2-5;和s图2)。首先,膈肌收缩的幅度,即Pes的负偏转相对较弱(≈-2.5 cm H2O),并且在所有组中相似;在“呼吸叠加”和“反向触发”中也观察到膈肌损伤,但在“辅助控制”中没有观察到。这表明在我们的研究中,膈肌损伤的机制不是剧烈的吸气努力,即辅助性肌肉损伤。其次,在机械呼吸到期期间,呼吸叠加和反向触发会产生膈肌收缩(图1)。因此,呼吸叠加和反向触发会导致膈肌的偏心收缩,导致膈肌损伤,即偏心性肌损伤。值得注意的是,偏心收缩比同心收缩更容易损伤肌肉,同心收缩是在肌肉缩短过程中产生的力。第三,快缩纤维(II型)比慢抽所纤维(I型)更容易受到偏心肌损伤。在我们的研究中,从不同步指数中可以明显看出,在呼吸叠加中观察到的偏心收缩比反向触发更频繁,这可能解释了呼吸堆叠与反向触发中肌纤维损伤的不同模式(见图2);第四,患有偏心收缩的膈肌高度参与细胞因子和趋化因子介导的促炎反应,支持我们的膈肌组织学分析。有趣的是,膈肌中的肌肉分化和生成在基因表达水平上受到抑制。这表明,偏心性膈肌损伤可能会抑制膈肌损伤后的肌肉再生,从而延长膈肌无力,从而导致困难脱机
五、研究局限性
本研究有几个局限性。首先,该模型(兔子,表面活性剂耗竭模型)是短期的,不能准确反映临床ARDS的较长正常时间过程。为了增强不同步对肺部和膈肌的影响,我们通过刺激双侧膈神经来模拟它,其不同步指数高于临床研究中的报道。因此,在将我们的结果外推到临床环境中时,需要谨慎。其次,膈神经刺激本身可能会影响膈功能,因为潜在的不应期、刺激增强和直接损伤。为了最大限度地降低这种风险,在整个4小时的方案中,在用冷盐水保护膈神经的同时,使用最小刺激强度来产生微弱的吸气力,即–2.5 cm H2O的Pes负偏转。膈肌产生的力在基线时是相似的 第三,呼吸叠加组吸气时间短,导致峰值流量(和压力)较高,这可能会影响肺损伤的差异。第四,预设的机械呼吸(95/min)和起搏呼吸(80/min)之间的频率差异可以实现反向触发。这导致随机相互作用,导致在不同时间点(起始和终止)出现各种反向触发表型(图1),反映了反向触发的临床场景。在我们的反向触发模型中,观察到反向触发和无效触发,其与呼吸频率的比例分别为47.8%±2%和8.2%±2%。因此,我们的模型在研究反向触发时是稳健的。在呼气期(而不是吸气期)发生的自发吸气努力被定义为无效触发,并在计算不同步指数时排除。
六、结论
呼吸叠加加重了肺损伤,增加了潮气量和吸气肺压力。呼吸叠加和反向触发(无呼吸叠加)导致膈肌损伤和功能障碍,可能是因为膈肌偏心收缩。因此,密切监测和管理患者与呼吸机的不同步性对于最大限度地减少ARDS患者自主呼吸对肺和膈肌的损伤可能很重要。
述评
急性呼吸窘迫综合征(ARDS)是临床常见的危重病,既往研究强表明,在机械通气过程中保留ARDS患者的自主呼吸是一把双刃剑。一方面,保留自主呼吸能够改善肺泡的通气,改善通气血流比例失调,保持膈肌活动,预防呼吸机诱导的膈肌功能障碍;另一方面,过强的自主呼吸出现人机不同步造成自我伤害性肺损伤,肺内炎症加重、膈肌向心收缩、膈肌无力。氧合功能恶化,增加病死率。不适宜的自主呼吸对机械通气的ARDS患者肺及膈肌损伤机制仍不明确,该研究提出呼吸叠加加重了肺损伤,增加了潮气量和吸气肺压力。呼吸叠加和反向触发导致膈肌损伤和功能障碍,可能原因为膈肌偏心收缩。密切监测和管理患者与呼吸机的不同步性对于最大限度地减少ARDS患者自主呼吸对肺和膈肌的损伤很重要。
编译:王志高
审校:王静静 陈霞霞
点评:郭峻氚
原始文献:
Hashimoto H, Yoshida T, Firstiogusran AMF, Taenaka H, Nukiwa R, Koyama Y, Uchiyama A, Fujino Y. Asynchrony Injures Lung and Diaphragm in Acute Respiratory Distress Syndrome. Crit Care Med. 2023 Jul 17. doi: 10.1097/CCM.0000000000005988. PMID: 37459198.
(仅供医学专业人士参考)
声明:古麻今醉公众号为舒医汇旗下,古麻今醉公众号所发表内容之知识产权为舒医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
不感兴趣
看过了
取消

 打赏
打赏
不感兴趣
看过了
取消
精彩评论
相关阅读




中国医院排行榜















 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612



©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:82736610
 京公网安备 11010802020745号
京公网安备 11010802020745号


