
除了EMT和血管生成之外,称为转移性生态位的特殊微环境对于播散癌细胞适应并在新组织中存活很重要。甚至在播散的癌细胞到达外周组织之前,原发性肿瘤就可以诱导转移性生态位,以促进其存活和生长。
整合素简介
整合素是一个跨膜细胞粘附分子大家族,由非共价α/β异二聚体组成(十八种α链和八种β链相互结合形成24种不同的异二聚体)。
可分为:精氨酸 - 甘氨酸 - 天冬氨酸(RGD)结合受体,白细胞特异性受体,层粘连蛋白受体和胶原受体等。
整合素结构 Cancer Cell. 2019
α和β亚基具有较长的胞外结构域和较短的胞质结构域,例外的情况包括白细胞整合素(αL、αM和αX)和胶原结合β1整合素的α亚基,它们在β螺旋桨结构域2和3之间插入一个I样结构域。
当存在时,I结构域参与配体与β亚基细胞外部分的I样结构域的结合。此外,β4整合素在结构上也是变异的,因为它具有一个大而独特的胞质结构域,具有两对III型纤维连接蛋白样重复序列,并连接到半桥粒的角蛋白,而不是肌动蛋白和细胞骨架。
变构驱动的双向信号传导 Cancer Cell. 2019
α亚基N末端的β螺旋与β亚基I样结构域和杂交域结合,形成配体结合袋和整合素的头部部分。
失活的整合素 表现出一种封闭的构象(“在膝盖处弯曲”):配体结合袋对配体的亲和力较低,面向质膜和“腿”(α亚基Calf-1和-2;β亚基I-EGF3、I-EGF4和膜近端尾部结构域βTD),跨膜区和细胞质结构域毗邻。
Talin与β亚基胞质结构域的结合引发了巨大的构象变化,包括“腿”的延伸和跨膜和胞质结构域的分离。
配体与部分活性的整合素结合 可以诱导相同的构象变化,增强整合素的激活。
活化整合素 被扩展,并具有一个配体结合能力的头部,暴露在细胞外空间,一个β细胞质尾与talin结合。这些机制确保了talin结合诱导整合素激活,相反,配体结合诱导talin结合,从而与肌动蛋白细胞骨架结合。
整合素的激活、聚簇和自主信号传导 Cancer Cell. 2019
目前研究表明,GTP-Rap1诱导MRL蛋白(如RIAM和lamellipodin),激活talin,后者反过来与整合素β亚基胞质结构域结合,将其从α亚基的相应部分分离出来。
Talin的结合因此触发了巨大的构象变化,从而导致整合素的激活。talin和kindlin都可以连接到肌动蛋白的细胞骨架,促进整合素的聚簇,启动强大的细胞内信号传导。
整合素信号除了组织细胞骨架从而影响心肌素相关转录因子(myocardin-related transcription factor,MRTF)/血清应答因子(serum response factor,SRF)外,整合素信号还有助于AP-1的激活、FOXO的抑制和YAP/TEAD的激活。整合素主要通过激活MRTF、c-Jun和YAP来影响这些转录因子,而受体酪氨酸激酶通过Ras-ERK和PI3K-AKT通路有效激活c-Fos并抑制FOXO。
整合素与炎症
在被激活时,整合素与其配体相互作用,介导白细胞的滚动、粘附、爬行和跨内膜迁移,以便经历组织归巢和炎症。整合素功能的适当调节对于控制炎症反应至关重要。
为了归巢于炎症部位,白细胞通过与内皮表面的选择素和趋化因子相互作用在血管内皮上滚动。这些相互作用触发由内而外和由外而内的信号级联反应,导致白细胞上的整合素激活成高亲和构象,促进整合素与其配位配体的结合(例如,αLβ2/ICAM-1; α4β1/ VCAM-1)和白细胞与内皮壁的牢固粘附。
此后,白细胞在称为爬行的过程中沿着内皮表面缓慢移动,这确保了所述白细胞的适当外渗位点的位置。爬行主要由整合素αLβ2或αMβ2介导,具体取决于白细胞亚型。随后,整合素αLβ2、αMβ2 和 α3β1 与各种配体相互作用,包括 ICAM-1/2 和连接粘附分子,以促进 白细胞的跨内皮迁移和分离。
Front. Cell Dev. Biol. 2020
整合素在适应性免疫中起重要作用。除了T细胞受体(TCR)和肽-MHC复合物之间的经典相互作用,抗原呈递细胞在称为免疫突触的特殊细胞界面上的成功抗原呈递和T细胞活化需要细胞的牢固粘附。在免疫突触中,αLβ2与ICAM-1的相互作用建立了一个独特的边缘区域,称为pSMAC;TCR和辅助分子在cSMAC中富集,这可能使T细胞被完全激活。
急性炎症反应的消退对于维持组织稳态至关重要,并有助于预防异常的慢性炎症。这主要是通过吞噬细胞在称为胞葬作用去除凋亡细胞(主要是中性粒细胞)来实现的,其中整合素是关键参与者。此外, 凋亡免疫细胞的清除将吞噬细胞重新编程为促消退表型。
整合素αVβ3和αVβ5在髓系细胞(分别为巨噬细胞和树突状细胞)上的表达可提高视凋细胞的清除率,而αVβ8以TGF-β1依赖的方式诱导调节性T(Treg)细胞的扩增。
这些αV的同时丢失,从而引发炎症性肠病(IBD)和自身免疫的发展。
在LPS诱导的肺部炎症模型中,凋亡细胞的气管内滴注被证明可以通过 CD11c+CD103+DC引发这些细胞的吞噬作用,从而启动 Treg 细胞的扩增并减轻肺部炎症。这些 DC 丢失整合素αV 导致凋亡细胞吞噬作用受损,抑制 TGF-β1 的产生和 Treg 细胞扩增,从而导致肺部炎症加重。
αV整合素是多个器官纤维化发作的主要参与者,但已经出现αV整合素的成纤维细胞特异性丢失,可能调节局部炎症反应。事实上,已经证明成纤维细胞特异性缺失αV整合素可减少17型驱动(但不是2型驱动)的肝和肺纤维化。这些纤维化模型不仅进一步加速了2型炎症标志物(IL-13)的同时增加,而且还导致成纤维细胞上缺乏α V的小鼠的肺和肝脏中嗜酸性粒细胞的积累。
细胞外基质僵硬和永久沉降是纤维化的特征,其涉及整合素信号传导和基质金属蛋白酶(MMP)介导的ECM降解的失调。
在慢性炎症条件下,间质细胞释放TGF-β1 和 IL-13 ,上调 细胞外基质的产生,导致发炎组织的纤维化进展。
整合素β1 也被证明可以调节肺部炎症,因为从2型肺泡上皮细胞(AECs)中删除β1会导致肺气肿和成年小鼠肺部巨噬细胞激增。
整合素与癌症
整合素几乎参与癌症进展的所有步骤,包括癌症的发生和增殖,局部侵袭和内渗到脉管系统,循环肿瘤细胞的存活,转移性生态位的启动,外渗到继发部位和新组织的转移性克隆。
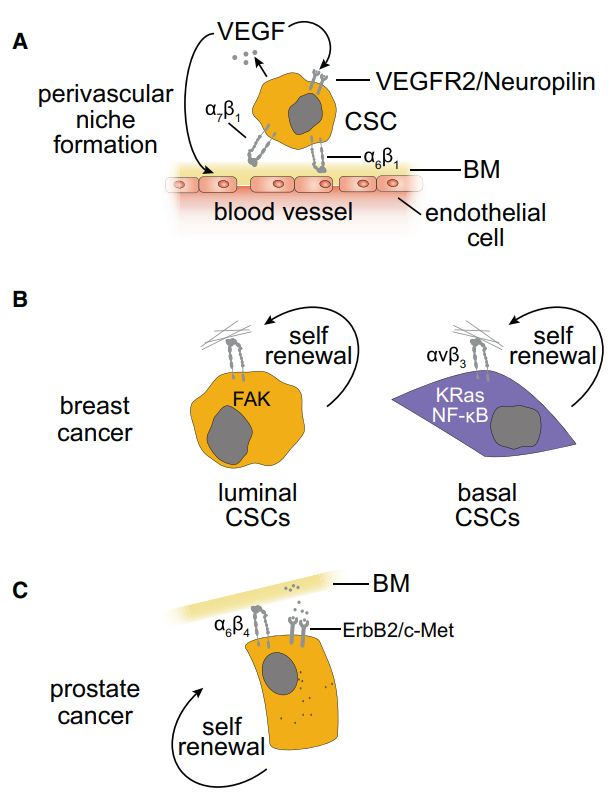
α3β1、α4β1、α5β1、α6β4、αVβ3、αVβ5、αVβ6 和 αVβ8 的表达被认为与转移和患者 预后不良 相关。
尽管癌细胞和微环境之间的整合素调控相当复杂,特别是癌症转移。
整合素与肿瘤转移 Cancer Cell 2019
特异性层粘连蛋白(LAM)亚型的分泌通过激活β1整合素信号通路促进转移部位癌的起始和重新启动。腱生蛋白c(Tenascin C,TNC)和骨膜蛋白(periostin,POSTN)也是促进转移再活化的关键基质元素。
基质元素和癌细胞通过包括中性粒细胞细胞外捕网(NET)重构在内的多种机制协同沉积转移性生态位的细胞外基质。LAM、纤维连接素(FN)、POSTN和TNC通过整合素、Syndecan-4(SDC4)和Wnt 受体(Frizzled,FZD)作用,激活参与转移生长的信号通路。
上皮-间充质转化(EMT)是癌症侵袭和转移的第一步 。EMT 通过下调蛋白质(如 E-钙粘蛋白)来丧失上皮特征,并通过表达间充质蛋白(α-SMA)、MMP 和增强的运动性而向成纤维细胞样表型转变。
整合素 在诱导EMT和介导其某些作用方面起着重要作用。主要的EMT诱导剂之一是TGF-β,它是一种抗炎和促进纤维化活性的细胞因子。
TGF-β先以失活前体分泌到细胞外空间,成熟的TGF-β被笼在潜伏期相关蛋白(latency-associated protein,LAP)中,从而限制其生物活性。
LAP 包含一个 整合素结合 的 RGD 基序,允许 αV 整合素结合并施加机械力来打开 LAP 笼。
在与受体结合后,成熟的TGF-β诱导上皮蛋白(如E-钙粘蛋白)的下调和间充质蛋白(如N-钙粘蛋白)的上调。
整合素α3β1表达 ,是 TGF-β刺激SMAD所需要的 , 用TGF-β刺激上皮细胞导致 β4整合素 (上皮完整性和稳定性所必需的典型上皮整合素)下调,从而促进迁移。
胶原蛋白交联和ECM固化加强了β1整合素聚簇和信号传导以及粘连。此外,由肿瘤相关成纤维细胞增加的层粘连蛋白链提供了改变的ECM沉积,有利于α6β4介导的癌细胞迁移。ECM重塑和降解与整合素功能相结合,对侵袭至关重要。
MMP 是胶原蛋白、纤连蛋白和层粘连蛋白的蛋白水解酶,是 ECM 的主要成分。在宫颈癌细胞系中,发现整合素α5β1-纤连蛋白相互作用可诱导 proMMP9 的表达和激活,并中度改变涉及 FAK、整合素连接的蛋白激酶、ERK、磷酸肌醇-3-激酶 (PI3K) 和 NF-κ B 信号传导级联。
整合素α5β1与层粘连蛋白的相互作用也诱导MMP-9表达并激活信号级联。还发现这种配体-整合素相互作用可以加速细胞迁移。此外,不是整合素α5β1,而且整合素αVβ3都与MMP活性有关。例如,整合素αVβ3表达是乳腺癌细胞骨转移的主要决定因素。此外,整合素αVβ3被发现是MMP-2活化和淋巴结转移的调节剂。
血管生成是癌症进展和转移的另一个重要因素。一般来说,缺乏血液循环的肿瘤在直径到1-2mm3然后停止;然而,在血管生成产生的区域时,它们可以生长 >2mm3。
在没有血管支持的情况下,由于氧气和营养供应不足,肿瘤可能会坏死甚至凋亡。血管生成不仅为肿瘤生长提供营养,还支持肿瘤细胞进入循环的逃生途径。
血管生成, 三种内皮整合素起着至关重要的作用:αVβ3、αVβ5 和 α5β1 。VEGF介导的血管生成需要αVβ5,FGF介导的血管生成需要αVβ3和α5β1发生。
在临床前研究中,用αVβ3拮抗剂抑制血管生成抑制肿瘤进展。
除了EMT和血管生成之外,称为转移性生态位的特殊微环境对于播散癌细胞适应并在新组织中存活很重要。甚至在播散的癌细胞到达外周组织之前,原发性肿瘤就可以诱导转移性生态位,以促进其存活和生长。
表达高水平的 αVβ5 的乳腺癌外泌体被证明播散到含有富含纤连蛋白的 ECM 的肝组织,而那些α6β4 的高水平外泌体播散到含有富含层粘连蛋白的 ECM 的肺组织。所提出的转移前生态位形成机制涉及肺和肝组织中发现的原炎性S100基因的激活。
参考资料
Park EJ, Myint PK, Ito A, Appiah MG, Darkwah S, Kawamoto E and Shimaoka M (2020) Integrin-Ligand Interactions in Inflammation, Cancer, and Metabolic Disease: Insights Into the Multifaceted Roles of an Emerging Ligand Irisin. Front. Cell Dev. Biol. 8:588066. doi: 10.3389/fcell.2020.588066
Jonathan Cooper, Filippo G Giancotti, Integrin Signaling in Cancer: Mechanotransduction,Stemness, Epithelial Plasticity,and Therapeutic Resistance,Cancer Cell. 2019 Mar 18;35(3):347-367.
Mezu-Ndubuisi OJ, et al. The role of integrins in inflammation and angiogenesis.Pediatr Res. 2021 May;89(7):1619-1626.
来源:闲谈 Immunology 2023-05-05
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
(京)网药械信息备字(2025)第00082号
京公网安备 11010802020745号
工商备案公示信息
(京)网药械信息备字(2025)第00082号

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您



































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




