
本研究表明,运动增强了这种沟通,运动对纹状体的影响是由肠道代谢产物决定的,此外,运动诱导的脊髓感觉神经元的活动通过MAO的调节驱动纹状体多巴胺信号。
编译:微科盟听雪斋,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读 运动对健康的生理学有着广泛的有益影响。然而,调节个人参与体育活动动机的机制仍不完全清楚。刺激参与竞技性和娱乐性运动的一个重要因素是长期体育活动所带来的愉悦感,这是由运动引起的大脑神经化学变化所触发的。本研究报告了在小鼠体内发现的一种肠-脑连接,它通过增强身体活动期间的多巴胺信号来增强运动表现。
结果发现,肠道中内源性大麻素代谢产物的微生物组依赖性产生刺激了表达TRPV1的感觉神经元的活动,从而在运动期间提高腹侧纹状体中的多巴胺水平。刺激这一通路可提高跑步能力,而微生物组耗竭、外周内源性大麻素受体抑制、脊髓传入神经元消融或多巴胺阻断则会削弱运动能力。这些结果表明,运动的奖赏特性受到肠道源性内感受回路的影响,并为运动表现的个体间差异提供了微生物依赖性解释。
论文ID
原名:A microbiome-dependent gut–brain pathway regulates motivation for exercise
译名:微生物组依赖性肠-脑通路调节运动动机
期刊:Nature
IF:69.504
发表时间:2022.12
通讯作者:Christoph A. Thaiss
通讯作者单位:美国宾夕法尼亚大学
DOI号:10.1038/s41586-022-05525-z
实验设计
结果
1 宏基因组对运动表现的影响
为了确定运动表现的新调节因子,我们使用了199只多样性离系繁殖(DO)小鼠,这些小鼠的基因组来源于8个明确的交叉遗传背景。我们通过单核苷酸多态性基因分型、血清代谢组学、粪便样本16S核糖体DNA(rDNA)测序和多参数代谢分析(图1a)对这一队列进行了深入分析,结果每只小鼠收集了10500多个数据点,总计接近210万个数据点。随后,小鼠通过自愿在轮子上跑步或在跑步机上耐力跑步来进行运动分析。所有小鼠在运动前均未接受训练。性能数据显示了动物在跑步机和轮式跑步中的显著差异(图1b,c和扩展数据图1a,b)。 为了探究这种差异背后的因素,我们首先关注宿主基因组。为此,我们对小鼠进行了基因分型,构建了遗传亲缘关系矩阵(扩展数据图1c),对每个跑步机参数进行了全基因组关联研究(扩展数据表1d),并计算了狭义遗传力。
对于每个终点,遗传力都相当低(低于0.2,扩展数据图1e),与之前的估计一致,这表明遗传对运动能力的个体间变异的贡献较小。 因此,我们转向了非遗传参数,并评估了它们对运动表现的相对贡献。血清代谢组、肠道微生物组组成和代谢参数在动物队列中差异很大(扩展数据图1f–o和补充表1)。我们使用机器学习方法识别对运动表现有很强预测作用的变量,再次使用跑步机参数作为终点。我们用所有可用的特征训练了一个梯度增强决策树算法来预测跑步机性能(图1d和扩展数据图1p,q)。为了确定驱动这些预测的特征,我们将所有变量分类为“metagroups”,并确定其相对贡献(扩展数据图1r)。值得注意的是,仅基于16S rDNA测序结果的预测获得的准确度几乎与血清代谢组或所有测量参数组合获得的准确度一样高,尽管包含的变量数量要少得多(图1e和扩展数据图1s–w)。相比之下,主要代谢和行为特征与耐力无关(扩展数据图1x–z)。 最近的报告强调了肠道微生物组在影响运动表现方面的潜在作用,但其总体重要性和潜在机制仍不清楚。为了确定DO小鼠运动表现的个体间差异在多大程度上是由微生物组驱动的,我们进行了功能丧失(微生物组耗竭)和功能获得(微生物组移植)实验(图1f)。使用广谱抗生素消除微生物群会降低转轮(图1g和扩展数据图2a)和跑步机(图1h和扩展数据表2b–d)上的运动表现。将DO小鼠的微生物组样本转移到无菌小鼠,导致供体和受体的转轮表现之间存在很强的相关性(图1i)。这些结果表明,微生物组对DO小鼠的运动能力有很强的功能性贡献。 为了确定微生物组中推动其对身体表现贡献的分类元素,我们转向了等基因C57BL/6J小鼠,这使得我们能够仅改变微生物组参数,同时保持所有其他因素不变。首先用多种正交方法测试了微生物组的因果关系。
广谱抗生素消除微生物组使跑步机和转轮的运动表现降低了约50%(图2a,b和扩展数据图2e–i)。为了确定这些观察结果不是由于口服抗生素的潜在副作用,我们在无菌条件下对无菌小鼠进行了运动试验,结果显示出了类似的性能下降(图2c,d和扩展数据图3j–n)。在雄性和雌性小鼠中观察到最佳运动表现的微生物组依赖性(扩展数据图2e–i,o–s)。重要的是,停止抗生素治疗以及无菌小鼠的常规化完全恢复了运动表现(扩展数据图2t–aa),表明微生物组消融对运动的影响是急性和可逆的。微生物组耗竭的影响仅局限于运动,一般不会影响运动,因为在抗生素治疗的小鼠中,居住笼和户外运动都没有受到损害(扩展数据图3a–e)。 为了缩小与表型相关的微生物组的可能成员,我们接下来减少了我们使用的抗生素组合的范围。仅使用可吸收或不可吸收性抗生素治疗同样减少了跑步机跑步(扩展数据图3f–i),进一步排除了可能的全身副作用。当我们以不影响体重的剂量单独使用每种抗生素时(扩展数据图3j),除新霉素外,所有抗生素在跑步机和转轮设置中都降低了运动表现(图2e、f和扩展数据图3k–n)。一致地,用新霉素处理的供体的粪便微生物组样本(显示正常的跑步能力)重建的无菌小鼠达到了用完全定殖的常规动物的样本重建的那些小鼠的跑步机参数,而使用氨苄青霉素处理的供体的微生物组样本的无菌受体(其运动表现下降)表现更差(扩展数据图3o–s)。这些结果表明,微生物组的某些分类元素对于运动调节既必要又又充分。
为了确定此类分类元素,我们对接受新霉素和氨苄青霉素处理的小鼠进行了16S rDNA测序(扩展数据图3t和补充表1),并将结果与从DO队列中获得的结果进行了比较分析。我们使用SHapley Additive exPlanation(SHAP)评估了预测模型中单个微生物组特征的贡献。我们首先根据微生物组元组中的所有变量的平均绝对SHAP值对其进行了排名(扩展数据图1r),该值指定了丹毒丝菌科和毛螺菌科的几个成员作为运动表现的可能驱动因素(扩展数据表3u)。这与丹毒丝菌科对新霉素耐药但对氨苄青霉素敏感一致,因此与微生物组转移至无菌小鼠后的运动活性呈正相关(扩展数据图3v,w)。然后,我们通过在无菌小鼠中进行单定殖实验来测试单个物种的贡献。我们用一种有助于预测运动表现的丹毒丝菌科成员(Eubacterium rectale)、一种同样参与预测运动的毛螺菌科成员(Coprococcus eutactus),一种对运动没有预测价值的毛螺菌科成员(Blautia producta)和一种分类上独特的微生物群成员(Ruminococcus gnavus)。我们通过16S定量PCR(qPCR)和测序验证单定殖成功(扩展数据图3x,y和补充表1),然后在跑步机上锻炼受体小鼠。与无菌小鼠相比,Eubacterium和Coprococcus提高了运动表现,达到了与常规定殖小鼠相当的水平,而Blautia和Ruminococcus则没有(图2g,h和扩展数据图3z,aa)。总之,这些数据证明了微生物组在调节运动表现中的致病作用,并强调了丹毒丝菌科和毛螺菌科的某些成员可能是分类贡献者。
图1. 肠道微生物组对小鼠运动表现的影响。a,对DO小鼠进行基因组、微生物组、代谢组和能量代谢分析,并评估其运动表现。b,c,跑步机(b)和转轮(c)性能参数的多样性。d,e,基于包括所有评估的非遗传特征(d)和仅来自16S rDNA测序的特征(e)的模型的算法预测与测量的跑步机距离。f,DO小鼠微生物组因果关系测试干预示意图。g,h,用抗生素(Abx)处理的DO小鼠的转轮(g)和跑步机(h)运动。i,微生物组从DO小鼠转移到无菌小鼠的供体和受体在跑步机上跑步的相关性。误差条表示平均值±s.e.m。**P<0.01;***P<0.001。
图2. 微生物群成员对运动表现的贡献。a,广谱抗生素(Abx)处理小鼠和对照组在跑步机上的距离的Kaplan–Meier图。b,Abx小鼠和对照组自主转轮运动的平均小时距离。插图显示了代表性的记录痕迹。c,无菌(GF)小鼠和对照组在跑步机上的距离的Kaplan–Meier图。d,GF小鼠和对照组自主转轮运动的平均小时距离。插图显示了代表性的记录痕迹。e,f,用指定抗生素处理的小鼠在跑步机上的距离的Kaplan–Meier图(e)和量化(f)。g,h,SPF小鼠、GF小鼠和用指定细菌物种单定殖的GF小鼠在跑步机上的距离的Kaplan–Meier图(g)和量化(h)。误差条表示平均值±s.e.m。NS,不显著;*P<0.05;**P<0.01;***P<0.0001。
2 微生物组对纹状体多巴胺的影响
作者接下来试图确定微生物组影响运动表现的机制。根据先前的报告,我们假设,最佳肌肉功能需要完整的微生物组。事实上,我们注意到抗生素处理小鼠的比目鱼肌、腓肠肌和胫前肌的质量减少(扩展数据图4a–c),而趾长伸肌没有受到影响(扩展数据表4d)。然而,肌肉功能、耗氧量或转录组学的体外测试并未显示抗生素治疗的主要影响(扩展数据图4e–s),这表明肠道微生物组对自愿运动和耐力锻炼的显著影响可能不是由肌肉生理学变化介导的。
除了肌肉骨骼功能外,运动表现的一个主要因素是动机状态。纹状体是大脑中与动机行为和身体活动启动密切相关的一个区域。为了探讨微生物组对运动时纹状体反应的影响,我们对接受抗生素处理的小鼠在跑步机运动前后的纹状体进行了单核RNA测序(RNA-seq)。我们对总共44100个细胞进行了测序,并基于标准标记物鉴定了细胞类型(扩展数据图5a–h)。然后,我们关注神经元群,并确定了表达多巴胺受体D1和D2的主要中棘神经元群(图3a和扩展数据图5i)。我们注意到,立早基因(如Arc和Fos)的表达因运动而显著升高;然而,在缺少微生物组的情况下,这种升高被抑制(图3b,c和扩展数据图5j,k)。这些数据表明,肠道微生物定殖对运动触发的纹状体激活很重要。
鉴于纹状体中棘神经元由多巴胺控制,而多巴胺是身体活动驱动的关键调节因子,我们接下来测量了微生物组耗竭小鼠的纹状体多巴胺水平。正如预期的那样,运动后纹状体多巴胺水平增加。值得注意的是,在微生物组耗竭的小鼠中,这种升高被减弱(图3d),表明肠道微生物定殖是运动后多巴胺有效释放所必需的。这不是因为微生物组缺失小鼠的运动能力降低,因为当第一只动物耗尽之前终止运动方案时,多巴胺反应也同样降低(图3d)。通过使用荧光多巴胺传感器实时测量多巴胺水平(图3e和扩展数据图6a),显示了不同运动时间后腹侧和背侧纹状体多巴胺的快速持续上调(扩展数据图6b-d)。引人注目的是,在抗生素处理的动物中,这种多巴胺反应几乎被消除(图3f–h),进一步强调了微生物组对纹状体多巴胺对身体活动反应的重要性。仅在运动状态下观察到纹状体多巴胺反应受损,而多巴胺的基础水平不受微生物组的影响(图3d),这与抗生素处理小鼠的完整基础运动一致(扩展数据图3a–e)。其他主要的纹状体神经递质,如谷氨酸和乙酰胆碱,既不受运动的影响,也不受微生物组的影响(扩展数据图6e,f)。
接下来,我们评估了运动引发的多巴胺反应是否在机制上参与了微生物组的表现刺激效应。首先,证据来自DO小鼠运动后纹状体多巴胺水平与转轮运动相关的发现(图3i)。其次,接受运动增强微生物组移植的无菌小鼠(无论是来自表现最好的DO小鼠还是来自常规定殖的C57BL/6小鼠)也显示出多巴胺水平升高(图3j和扩展数据图6g)。
第三,通过药理学多巴胺受体拮抗剂舒必利或通过对腹侧被盖区(VTA)多巴胺能神经元的化学遗传抑制来抑制多巴胺信号传导,都反映了微生物组耗竭的影响(图3k,l和扩展数据图6h–m)。此外,药理学多巴胺受体激动剂溴隐亭对多巴胺信号的激活恢复了抗生素处理小鼠的运动表现(图3m,n)。这些实验表明,纹状体多巴胺信号是微生物组影响运动活动的重要媒介。
因此,我们试图探索微生物组消融损害纹状体中运动诱导的多巴胺水平的机制。鉴于多巴胺能VTA神经元在纹状体活动调节中的重要性,我们通过在运动期间使用钙成像来监测其活动动态(扩展数据图6n)。然而,我们没有观察到任何抗生素诱导的钙通量差异(扩展数据图6o–r),这使得微生物组对多巴胺水平和身体表现的影响不太可能由VTA神经元活性的差异介导。一致地,瘦素的施用(已证明通过VTA13抑制运动的奖赏效应)与运动抑制中的微生物组消融具有叠加效应(扩展数据图7a–d)。我们接下来考虑了多巴胺水平可能受到突触后纹状体不同转换的影响的可能性。多巴胺降解由单胺氧化酶(MAO)调节。值得注意的是,在运动小鼠中,Maoa的纹状体表达受到抑制,这在缺乏肠道微生物组的情况下不会发生(图3o)。因此,我们假设,在微生物组耗竭小鼠中,持续的MAO水平是多巴胺对运动反应迟钝的原因,阻断MAO会使多巴胺信号正常化。事实上,在抗生素处理的小鼠中,药理学MAO抑制增强了运动诱导的多巴胺水平,并恢复了跑步表现(图3p–r和扩展数据图7e–h)。这些结果表明,肠道微生物定殖通过限制MAO驱动的多巴胺周转来增强纹状体中运动诱导的多巴胺信号传导。
图3. 微生物组影响纹状体中运动诱导的多巴胺反应。a,运动前和运动期间对照组和Abx处理小鼠纹状体单核RNA-seq神经元的UMAP图。b,c,运动前和运动期间对照和Abx处理小鼠的纹状体神经元中Arc表达的UMAP图(b)和定量(c)。d,Abx处理小鼠和对照组在稳定状态下、耐力运动后和定时运动60分钟后大脑中的多巴胺水平。e–h,示意图(e),注射到伏隔核的多巴胺传感器的纤维光度记录的记录痕迹(f)、量化(g)和平均信号(h)。在运动方案结束时记录小鼠。I-j,DO小鼠(i)和DO小鼠微生物群样本的无菌受体(j)在自愿运动中纹状体多巴胺浓度和跑步距离的相关性。k–n,注射舒必利(k,l)或溴隐亭(m,n)的Abx处理小鼠的自主转轮运动的平均小时距离(k,m)和量化(l,n)。插图显示了代表性的记录痕迹。o,对照组和Abx处理小鼠运动前后纹状体中Maoa的表达。p,在稳定状态和运动后,腹腔注射赋形剂或MAO抑制剂pargyline,Abx处理组和对照组小鼠大脑中的多巴胺水平。q-r,注射MAO抑制剂pargyline的Abx处理小鼠的自主转轮运动的平均小时距离(q)和定量(r)。插图显示了代表性的记录痕迹。误差条表示平均值±s.e.m。*,P<0.05;**,P<0.01;***,P<0.001;***,P<0.0001。
3 感觉神经元对运动的调节
接下来,我们探究肠道微生物定殖如何影响中脑多巴胺水平。从肠腔到大脑的信号转导可以通过循环因子(如微生物代谢产物)或通过传入神经元回路进行。我们首先关注体液因素,因为我们在抗生素处理小鼠中观察到血清代谢物谱显著改变(扩展数据图7i)。然而,DO队列中微生物组依赖性代谢产物和运动相关代谢产物之间的相关性较弱,没有产生任何强大的候选体液介质(扩展数据图7j)。 因此,我们关注肠道神经支配的传入感觉神经元。我们使用了失神经Trpv1DTA小鼠,其缺乏大量表达香草素受体TRPV1的脊髓和迷走神经传入神经元。值得注意的是,在转轮和跑步机上,Trpv1DTA小鼠的运动能力受损的方式与微生物组耗竭小鼠相似,抗生素处理并未进一步调节其运动表现(图4a–d和扩展数据图7k,l)。在表达TRPV1的神经元的化学遗传学沉默后,观察到了类似的运动表现受损(扩展数据图7m–p)。使用低剂量TRPV1激动剂辣椒素激活TRPV1+传入神经元,恢复了抗生素处理小鼠的转轮和跑步机运动(图4e,f和扩展数据图7q–t)。 鉴于TRPV1广泛由感觉神经元表达,我们试图确定脊髓或迷走神经传入是否是微生物组影响运动的主要媒介。
为此,我们检查了运动小鼠的背根神经节(DRG)和结节神经节(NG)。我们注意到,如cFos所示,跑步机运动诱导TRPV1+神经元的神经元活动(扩展数据图8a–c)。抗生素处理减少了DRG中运动诱导的神经元激活,但不影响NGs(图4g和扩展数据图8d–i),表明脊髓通路对微生物组衍生信号更敏感。一致地,NG注射CCK–SAP对肠神经支配的迷走神经传入的消融不会影响运动能力(扩展数据图9a,b),而通过手术切除腹腔/肠系膜上神经节(CSMG)切断肠神经支配的脊髓传入纤维会降低运动表现,类似于微生物组消融(扩展数据图9c-j)。
为了探讨感觉神经元的刺激是否影响MAO-多巴胺通路,测量了纹状体中的Maoa转录本和运动后多巴胺水平。我们发现辣椒素处理恢复了微生物组缺失小鼠的Maoa转录本和正常多巴胺水平,而Trpv1DTA小鼠的运动后多巴胺水平与缺乏完整微生物组的小鼠一样低(图4h,i)。类似地,CSMG的去除提高了Maoa的表达并抑制了多巴胺水平(扩展数据图9k,l)。腹侧纹状体伏隔核中多巴胺传感器的实时记录证实,运动后纹状体多巴胺水平通过辣椒素的外周给药得以恢复(图4j–l和扩展数据图9m)。为了确定辣椒素的运动刺激作用是否在功能上依赖于多巴胺信号,我们首先在单个动物的水平上比较了纹状体多巴胺水平与运动表现,并发现了高相关性(扩展数据图9n)。然后,我们用辣椒素和舒必利共同处理微生物组耗竭的小鼠和对照小鼠,在刺激传入神经元的同时抑制多巴胺信号。重要的是,舒必利治疗阻断了辣椒素的运动增强作用(图4,n),表明多巴胺信号确实是必需的。此外,溴隐亭对多巴胺受体的刺激绕过了对微生物组和传入感觉神经元的需求,并完全恢复了Trpv1DTA和抗生素处理小鼠的转轮运动(图4o,p)。总之,这些结果表明,微生物组对纹状体多巴胺水平和运动表现的影响需要TRPV1+神经元。
图4. 微生物组通过传入感觉神经元影响运动表现。a、b,野生型(WT)和Trpv1DTA小鼠在接受和不接受Abx处理的情况下的自愿转轮运动的平均小时距离(a)和定量(b)。插图显示了代表性的记录痕迹。c、d,对照组、Abx处理小鼠和Trpv1DTA小鼠在跑步机上的距离的Kaplan–Meier图(c)和量化(d)。e、f,Abx处理小鼠和对照组在有或没有辣椒素治疗的情况下的跑步机距离的Kaplan–Meier图(e)和量化(f)。g,无论是否使用Abx处理,久坐和运动后小鼠DRG中Arc的表达。h,运动后服用或不服用辣椒素的Abx处理小鼠和对照小鼠纹状体中Maoa的表达。i,对照小鼠、Abx处理小鼠和Trpv1DTA小鼠(无论是否接受辣椒素治疗)大脑中的多巴胺水平。j–l,记录运动后Abx处理小鼠和对照组(有或没有辣椒素治疗)的痕迹(j)、量化(k)和最大信号(l),无论是否使用辣椒素治疗。m、n,接受或不接受辣椒素和舒必利处理的Abx处理小鼠和对照组的自愿转轮运动的平均小时距离(m)和定量(n)。o、p,使用或不使用溴隐亭治疗的Abx处理和Trpv1DTA小鼠的自愿转轮运动的平均小时距离(o)和定量(p)。插图显示了代表性的记录痕迹。误差条表示平均值±s.e.m。*P<0.05;**P<0.01;***P<0.001;****P<0.0001。
4 肠道代谢物对运动的调节
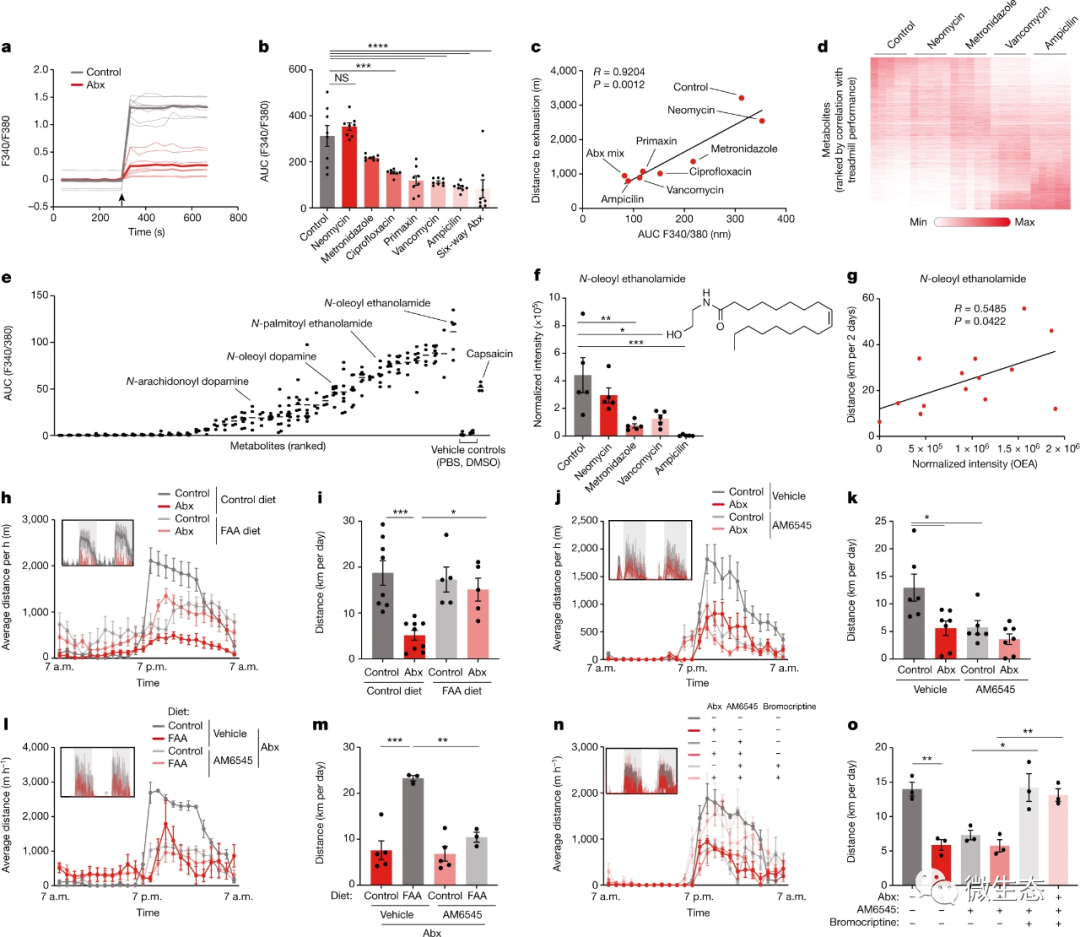
接下来,研究了微生物组影响肠道感觉神经活动以影响运动表现的可能机制。为了模拟传入神经元对管腔成分的感知,在体外将分离的DRG神经元暴露于来自抗生素处理小鼠、无菌小鼠和常规定殖对照的粪便提取物,并通过钙成像记录其活性。微生物组缺失小鼠提取物中的刺激电位降低(图5a和扩展数据图9o),表明DRG神经元可能感知到一种管腔成分。类似地,与定殖对照组相比,无菌小鼠的管腔含量显示DRG神经元刺激减少,这在无菌小鼠常规化时是可逆的(扩展数据图9p,q)。为了探究微生物组产物对DRG神经元的刺激是否与纹状体多巴胺水平和运动表现相关,我们使用了DO队列的粪便提取物,并测试了其DRG神经元刺激能力。观察到广泛的钙信号,这与纹状体中活动诱导的多巴胺释放和运动表现相关(扩展数据图9r–u)。 为了确定刺激DRG神经元的特定微生物组成分,我们再次使用了我们的差异抗生素治疗策略。我们首先将DRG神经元暴露于使用不同抗生素的小鼠粪便提取物,并观察到钙信号与动物的运动表现密切相关(图5b、c和扩展数据图9v)。然后,我们进行了非靶向代谢组学,并将结果与运动参数相关联(图5d)。之后筛选候选分子以激活DRG神经元中的钙信号(图5e和扩展数据图9w)。有趣的是,在最有效的代谢产物中有许多脂肪酸酰胺(FAAs),如N-油酰乙醇胺(OEA),其DRG刺激活性与辣椒素相当(图5e和扩展数据图9x,y)。在DO小鼠中,OEA水平被那些损害运动表现并与转轮运动相关的抗生素耗尽(图5f,g),这表明FAA代谢产物(如OEA)可能刺激感觉神经元以促进身体活动。 为了确定FAA是否足以在缺乏微生物组的情况下恢复运动能力,我们采用了膳食补充方法。我们选择了五种具有DRG刺激活性的FAAs(扩展数据图10a,b),并将它们补充到啮齿动物的饮食中(扩展数据表10c,d)。当喂食抗生素处理的小鼠时,这种FAA饮食恢复了粪便提取物诱导神经元钙信号的能力(扩展数据图10e,f)。引人注目的是,在没有微生物组的情况下,它还恢复了DRG活性、纹状体Maoa表达、多巴胺水平和运动表现(图5h,i和扩展数据图10g–l)。它同样增强了微生物组消融DO小鼠的跑步能力(扩展数据图10m–o)。鉴于OEA已被证明能在饮食参与的情况下刺激多巴胺的释放,我们探究胃内输注FAA是否足以概括运动对纹状体多巴胺水平的影响。在久坐的小鼠中,我们没有观察到多巴胺对FAA输注的反应。然而,在跑步机上运动时,肠道FAAs增强了身体活动诱导的多巴胺释放(扩展数据图10p,q)。
这些结果表明,肠道FAAs增强了运动的动机。 接下来,我们调查了微生物组产生的FAA是否能够解释其性能刺激效应。运动增强细菌单定殖无菌小鼠的OEA水平升高表明,FAA的微生物群产生可能会刺激跑步表现(扩展数据图11a)。有趣的是,最近发现某些共生微生物,包括来自丹毒丝菌科的Eubacterium和来自毛螺菌科的Coprococcus,含有负责FAA生物合成的生物合成基因簇。为了测试微生物组中该基因簇的存在是否足以增强运动,我们生成了一种含有该基因簇必需基因的大肠杆菌菌株,并用含有FAA生物合成基因簇或空载体的大肠杆菌菌株定殖无菌小鼠(扩展数据图11b)。我们首先验证了基因簇赋予大肠杆菌在体内产生FAA样OEA的能力,并刺激DRG神经元中的钙信号(扩展数据图11c–E)。然后,我们对单定殖小鼠进行了实验,以确定工程菌株对身体活动的影响。值得注意的是,虽然空载体对照菌株没有改善无菌小鼠的表现,但FAA生产菌株增强了小鼠在转轮和跑步机上的跑步能力(扩展数据图11f–k)。这些发现强调了能够产生代谢产物科的微生物基因簇可以影响宿主的运动能力
最后,我们探讨了肠道FAA刺激感觉神经元的机制及其对运动动机的影响。一些FAA代谢物充当内源性大麻素受体CB1的激动剂,该受体由TRPV1+DRG神经元表达(扩展数据图11l,m)。因此,这些观察结果提出了FAA刺激CB1可能会驱动感觉神经元活动、纹状体多巴胺释放和运动表现的可能性。与此假设一致,CB1缺陷小鼠表现出运动后DRG神经元活性降低、纹状体多巴胺减少和运动能力受损(扩展数据图11n–t),这与先前的发现一致。类似地,CB1受体拮抗剂O-2050抑制了转轮运动,而CB1受体激动剂CP55,940增强了表现(扩展数据图12a–d)。为了区分外周CB1信号传导对大脑的影响和内源性大麻素效应,我们使用了外周限制性CB1抑制剂AM6545,其复制了CB1缺乏的影响,并导致了与微生物组消融类似的运动损伤(图5j,k和扩展数据图12e–h)。重要的是,与AM6545联合治疗消除了增强运动的肠道干预措施的影响,如膳食补充FAA和用产生FAA的大肠杆菌单定殖,表明这些干预措施通过外周CB1信号传导起作用(图5l,m和扩展数据图12i–m)。我们想知道外周CB1刺激对运动的影响是否由纹状体中的多巴胺信号传导介导。事实上,AM6545处理提高了Maoa表达,并减弱了运动诱导的多巴胺激增(扩展数据图12n,o)。此外,AM6545处理的小鼠在接受溴隐亭多巴胺受体刺激时恢复了完全的运动能力(图5n,o),表明外周CB1信号对运动的影响是由多巴胺介导的。
图5. 外周内源性大麻素促进运动表现。a,记录暴露于Abx处理小鼠和对照组粪便滤液的DRG神经元的钙成像痕迹。箭头表示处理时间。b、c,暴露于用不同Abx处理的小鼠粪便滤液的DRG神经元的钙成像的量化(b)和与跑步机距离(c)的相关性。箭头表示处理时间。d,用不同Abx处理的小鼠盲肠内容物中检测到的代谢物热图。e,暴露于单个代谢物的DRG神经元的钙成像定量。f,用不同Abx处理的小鼠盲肠内容物中OEA的标准化丰度。g,DO小鼠OEA丰度与转轮运动的相关性。h–o,用或不用外周CB1拮抗剂AM6545(j–o)和多巴胺受体激动剂溴隐亭(n,o)治疗,喂食FAA补充或对照饮食(h,i,l,m)的Abx小鼠和对照组的自愿转轮运动的平均小时距离(h,j,l,n)和量化(i,k,m,o)。插图显示了代表性的记录痕迹。误差条表示平均值±s.e.m。*,P<0.05;**P<0.01;***P<0.001;****,P<0.0001。
讨论
遵守世界卫生组织的建议,每周至少进行150分钟的体育活动,将大大减轻全球疾病负担。但运动是剧烈的,除了心血管和呼吸健康外,还需要在职业、娱乐或治疗环境中保持强烈的动机状态。在这项研究中,证明了参与调节身体活动动机的大脑回路并非严格意义上的中枢神经系统自主,而是由源自肠道微生物群落的外围影响形成的,这为理解运动动机和表现的个体间差异提供了可能的机制基础。 通过整合深入剖析的、遗传和宏基因组多样的小鼠、非生物条件下的运动试验、神经生物学干预和代谢组学,以揭示肠道微生物组对参与运动表现的脑回路的影响。结果表明,肠道微生物组有助于肠道FAA代谢产物的产生,这些代谢产物触发CB1表达TRPV1+感觉神经元,进而向大脑发送运动诱导的传入信号,并促进纹状体MAO表达的下调。MAO的这种下调有助于提高多巴胺水平和增强运动能力(扩展数据图12p)。在缺少微生物组的情况下,传入信号被抑制,MAO水平被维持,因此运动诱发的多巴胺激增被减弱,对身体表现产生深远影响。 这些发现有几个重要的意义。首先,他们认为,“跑步高潮”背后的神经化学效应,即长时间体育活动后内源性大麻素释放所驱动的愉悦、奖励、焦虑和镇痛现象,可能受到胃肠道的影响。一致地,我们发现运动的镇痛作用同样依赖于肠道微生物定殖和外周CB1信号(扩展数据图12q,r),表明本研究中发现的途径可能调节运动生理的其他方面。从进化的角度来看,肠道代谢产物对运动诱导的奖赏和激励回路的调节可能有助于将营养物质的可用性和肠道微生物定殖状态与参与长时间体育活动的准备和能力相结合。其次,这些结果扩展了我们关于肠道和纹状体之间交流的神经通路的了解。
本研究表明,运动增强了这种沟通,运动对纹状体的影响是由肠道代谢产物决定的,此外,运动诱导的脊髓感觉神经元的活动通过MAO的调节驱动纹状体多巴胺信号。到目前为止,本研究还没有揭示感觉神经元活动调节纹状体中MAO表达的精确(多突触)回路。对神经元身份和控制前脑MAO水平的多突触回路的更详细的了解,将通过专注于胃肠道的非侵入性策略,提供对这种关键酶调节的更精确的处理。 最后,本研究表明,中边缘多巴胺依赖的奖赏和动机回路可以通过源自胃肠道的内感受通路进行调节。这一发现可能表明,其他依赖纹状体多巴胺信号的行为同样可以通过生活方式干预、饮食或代谢产物补充来改变。因此,它为更普遍的“interceptomics”概念铺平了道路,即刺激传入感觉通路的分子,从而通过外围干预影响大脑活动。如果适用于人类,本研究结果表明,刺激运动动机的interoceptomimetics可能会提供一个强有力的机会来抵消久坐生活方式对健康的有害影响。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




