
简单地说,肠道微生物及其代谢产物介导褪黑素对SD诱导的认知障碍的改善作用。
编译:微科盟煎蛋,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
睡眠不足是一个严重的全球健康问题,带来的后果包括记忆缺陷和胃肠道功能障碍。我们之前的研究表明,褪黑素可以有效改善睡眠剥夺(SD)引起的认知障碍和肠道微生物群紊乱。本研究进一步探讨了外源性褪黑素预防SD诱导的认知障碍的机制。本研究建立了粪便微生物群移植、气单胞菌定植和LPS或丁酸盐补充试验,以评估肠道微生物群及其代谢物在褪黑素缓解SD诱导的记忆障碍中的作用。结果表明,将SD肠道微生物群移植到正常小鼠中,诱导海马小胶质细胞过度激活和神经元凋亡、认知能力下降和结肠微生物群紊乱,表现为气单胞菌和LPS水平升高以及Lachnospiraceae_NK4A136和丁酸盐水平降低。所有这些事件都通过移植SD+褪黑素肠道微生物群后得到逆转。气单胞菌定植和LPS的添加在小鼠海马中产生炎症反应和空间记忆障碍。通过补充褪黑素,伴随着气单胞菌和LPS水平的降低,这些变化得以逆转。睡眠剥夺小鼠补充丁酸盐可恢复炎症反应和记忆障碍。在体外,补充LPS引起BV2细胞的炎症反应,通过补充丁酸盐得到改善。丁酸盐的这种改善作用被MCT1抑制剂和HDAC3激动剂预处理所阻断,但被TLR4和p-P65拮抗剂模拟。综上所述,肠道微生物及其代谢物介导褪黑素对SD诱导的认知障碍的改善作用。一种可行的机制是褪黑素下调气单胞菌和LPS的水平,并上调结肠中Lachnospiraceae_NK4A136和丁酸盐的水平。这些变化通过TLR4/NF-κB和MCT1/HDAC3信号通路之间的串扰减轻了海马中的炎症反应和神经元凋亡。
论文ID
原名:Gut microbiota-derived metabolites mediate the neuroprotective effect of melatonin in cognitive impairment induced by sleep deprivation
译名:肠道微生物群衍生代谢物介导褪黑素在睡眠剥夺引起的认知障碍中的神经保护作用
期刊:Microbiome
IF:16.837
发表时间:2023.1
通讯作者:陈耀星
通讯作者单位:中国农业大学动物医学院
DOI号:10.1186/s40168-022-01452-3
实验设计
结果
1 肠道菌群介导褪黑素在SD诱导的神经炎症和记忆障碍中的神经保护作用
为了测试Mel对SD诱导的记忆障碍的神经保护作用是否取决于肠道微生物群,我们将CON、SD或SD+Mel组的粪便微生物群移植到肠道微生物群耗尽的小鼠(经过10天的抗生素预处理)中2周(图2A)。抗生素的添加严重消耗了小鼠粪便中的细菌(图2B)。行为结果表明,SD-FMT组的潜伏期比CON-FMT组高115.8%(P = 0.002),而第一次测试(使用隐藏平台)的路径长度长143.3%(P = 0.002)(图2C,E-F)。各组之间的路径效率无显著差异(图2G)。在没有隐藏平台的第二个测试中(图2D),SD-FMT组进入目标区域的次数比CON-FMT组低55.2%(P = 0.005),而花费的时间比CON-FMT组低36.4%(P = 0.01)。因此,SD+Mel-FMT组和CON-FMT组之间没有显著差异(图2D,H,I)。与SD供体相比,携带来自SD+Mel供体微生物群的小鼠的记忆障碍有所改善,这表明肠道微生物群可能有助于Mel对SD小鼠认知功能障碍的改善(图2C-I)。
为了研究神经炎症是否由SD来源的肠道微生物群诱导,我们评估了海马中炎症因子和小胶质细胞免疫组化染色的变化。与CON-FMT组相比,SD-FMT组海马CA1、CA3和齿状回(DG)区域Iba1阳性细胞的累积光密度(IOD)分别高51.9% (P < 0.001)、27.6% (P = 0.01)和32.3% (P = 0.002)(图2J,K)。与CON-FMT小鼠相比,我们还观察到SD-FMT小鼠海马中IL-6 (36.5%,P = 0.03)和TNF-α (47.2%,P = 0.01)水平显著升高,IL-4 (49.6%,P = 0.04)和IL-10 (35.5%,P < 0.001)水平显著降低(图2L-O)。然而,“SD+Mel菌群”的FMT显著抑制了小胶质细胞的激活,增加了促炎因子,减少了抗炎因子。过度炎症可引起细胞毒性,干扰细胞生长,诱导细胞凋亡。因此,我们用蛋白质免疫印迹方法测定了cleaved caspase-3、Bax和Bcl-2的表达水平。与CON-FMT组相比,SD-FMT组Bcl-2表达明显下调(P = 0.021),cleaved caspase-3表达明显上调(P= 0.002),Bax表达明显上调(P = 0.001)。然而,“SD+Mel菌群”的FMT逆转了这些变化(补充图1)。FMT实验表明,肠道微生物群是Mel对SD诱导的神经炎症、细胞凋亡和记忆损伤的保护作用所必需的。
图1 实验模型示意图和时间轴。
A,粪便菌群移植(FMT)实验。B,Aeromonas veronii定植实验。C,LPS处理实验。D,丁酸盐处理实验。Mel:褪黑素,MWM: Morris水迷宫,Vehicle:2%乙醇无菌生理盐水。SD:睡眠剥夺,TAK-242: TLR4抑制剂。
图2 肠道菌群介导褪黑素在睡眠剥夺引起的记忆障碍中的神经保护作用。
A,实验设计示意图。B,对照组和ABS处理小鼠粪便中细菌集落形成单位(CFU)的比较(n= 10)。C,空间记忆测试轨迹图(带隐藏平台)。D,空间记忆测试轨迹图(无隐藏平台)。E,到达平台的潜伏期(n = 8)。F,到达平台的路径长度(n = 8)。G,到达平台的路径效率(n = 8)。H,停留在目标区域的时间(n = 8)。I,进入目标区域的次数(n = 8)。J,不同实验组的免疫组化小胶质细胞图像。免疫组化结果用ImageJ处理。比例尺= 50 μm。K,海马CA1、CA3和齿状回(DG)区域Iba1阳性细胞的IOD (n = 6)。L-O,海马中细胞因子(TNF-α、IL-6、IL-4和IL-10)的水平(n = 5)。CON-FMT:接受对照菌群FMT小鼠,SD-FMT:接受睡眠剥夺菌群FMT小鼠,SD+Mel-FMT:接受SD+Mel (20 mg/kg)菌群FMT小鼠,V-FMT:接受赋形剂菌群FMT小鼠。数据代表均值±SEM,* P < 0.05,** P < 0.01,*** P < 0.001。
2 FMT处理可调节受体小鼠肠道菌群组成
为了测试FMT是否调节肠道菌群,我们对受体小鼠进行了16S rDNA基因测序,以分析微生物处理后的细菌分类组成。从三组(n = 7)小鼠中共获得21个样本,随后测序生成V1-V9 16S rRNA基因谱。CON-FMT组、SD-FMT组和SD + Mel-FMT组总共获得了91308、91002和90919个原始reads。CON-FMT组、SD-FMT组和SD+Mel-FMT组分别有78824、77428和77628个clean tags。α多样性反映了微生物群的丰富性和多样性。3个处理组的Chao1、ACE、Simpson和Shannon指数均无明显变化(P > 0.05)(图3A,B,补充图2)。为了衡量微生物群落之间的相似程度,进一步使用Bray-Curtis PCoA评估β-多样性。结果显示各组间存在分离(图3D),主成分PC1、PC2和PC3分别解释了17.96%、15.50%和9.66%的变异(Adonis,P = 0.001,R2 = 0.379)。UPGMA结果显示,SD+Mel-FMT组比SD-FMT组更接近CON-FMT组,进一步验证了PCoA的结果(图3J,K)。在门水平上,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)和疣微菌门(Verrucomicrobiota)占优势(图4A)。在属水平上,uncultured_bacterium_f_Muribaculaceae、Lachnospiraceae_NK4A136_group、Akkermansia和乳杆菌属(Lactobacillus)是优势菌群(图4B)。
为了确定与CON-FMT、SD-FMT和SD+Mel-FMT组相关的特定细菌门,进行LDA和LEfSe分析以确定最有可能解释组间差异的核心类群。如图4C所示,与CON-FMT组和SD+Mel-FMT组相比,SD-FMT组中拟杆菌门和变形菌门丰度更高(拟杆菌门P = 0.046,LDA评分= 4.39;变形菌门P = 0.012,LDA评分= 3.98)。此外,如图4A所示,与CON-FMT组和SD+Mel-FMT组相比,SD-FMT组厚壁菌门数量减少(P < 0.05)。此外,LEfSe分析在三组中鉴定出64个分类生物标志物,LDA评分>3,P < 0.05。SD-FMT组Lachnospiraceae_nk4a136_group (P= 0.041,LDA评分= 4.55)、Eubacteriumxylanophilum_group(P= 0.027,LDA评分= 3.39)、Ruminococcus_1 (P = 0.018,LDA评分= 3.56)和Lachnospiraceae_A2 (P = 0.001,LDA评分= 3.64)的相对丰度显著低于CON-FMT组和SD+Mel-FMT组(图4G-J)。此外,SD-FMT组中Turicimonas的相对丰度(P = 0.035,LDA评分= 3.12)显著高于CON-FMT组和SD+Mel-FMT组(图4K),而CON-FMT组与SD+Mel-FMT组之间没有显著差异(P >0.05)。
图3 FMT处理小鼠结肠菌群的组成。
A、B,通过α多样性指数(Chao和Ace)评估结肠微生物丰富度和均匀度。C,主成分分析。D,PCoA评分图。E,基于肠道微生物OTU的binary_jaccard距离图的非度量多维尺度(NMDS)评分图。F,香农曲线。G,OTU等级曲线。H,稀疏曲线。I,丰度等级曲线。J,非加权配对算术平均法(UPGMA)(门水平)。K,CON-FMT组、SD-FMT组和SD+Mel-FMT组小鼠盲肠的非加权配对算术平均法(UPGMA)(属水平)分析。数据代表均值±SEM,*P < 0.05,**P < 0.01,***P < 0.001。
图4 FMT处理小鼠结肠菌群的组成和关键菌群。
A,各组结肠菌群在门水平上的相对丰度。B,各组肠道菌群在属水平上的相对丰度。C,采用LEfSe分析来确定不同组间有差异的细菌。D-K,基于LefSe结果的结肠菌群中p_Bacteroidota、p_Proteobacteria、c_Gammaproteobacteria、g_Lachnospiraceae_NK4A136、g_Eubacterium_xylanophilum、g_Ruminococcus_1、g_ Lachnospiraceae_A2和g_Turicionas的相对丰度。实线和虚线分别表示平均值和中值。CON-FMT:接受对照菌群FMT小鼠,SD-FMT:接受睡眠剥夺菌群FMT小鼠,SD+Mel-FMT:接受SD+Mel (20 mg/kg)菌群FMT小鼠。
3 FMT处理可调节受体小鼠肠道菌群代谢物组成
肠道菌群可以通过其代谢产物影响宿主。因此,我们对FMT受体小鼠进行了代谢组学分析。代谢物分析显示,结肠中有2260种代谢物。维恩图显示,不同的处理导致不同的代谢物变化(图5A)。PCA分析显示各组微生物群代谢产物组成明显聚类,与SD-FMT组相比,CON-FMT组更接近(图5B)。为了进一步验证不同组样本之间的差异,我们将正交投影应用于潜在结构判别分析(OPLS-DA)来实现这一目标。OPLS-DA模型显示三组间有良好的分离。预测OPLS-DA模型的质量参数值为[R2X(cum) = 0.587,R2Y(cum) = 0.474],适应度为[Q2(cum) = 0.273],模型具有良好的可靠性和可预测性(图5C,D)。火山图根据统计值(P < 0.05,|log2FC| > 1)显示差异代谢物的上调和下调(图5E-G)。具体而言,与CON-FMT组相比,SD-FMT组有547种代谢物上调,15种代谢物下调。但与SD-FMT组相比,SD+Mel-FMT组有574种代谢物增加,26种代谢物减少。此外,我们筛选了三组中41个变化最大的代谢物(图5J)。与SD-FMT组相比,SD+Mel-FMT组丁酸(P = 0.03)和L-色氨酸(P = 0.02)含量显著升高(图5H,I)。
图5 FMT处理小鼠结肠微生物代谢产物的组成。
A,基于微生物代谢产物的维恩图。B,β多样性主成分分析(PCA)。C,对结肠进行潜在结构判别分析(OPLS-DA)的正交投影评分图。D,在排列验证图中,R2和Q2的y轴截距分别为0.587和0.273,说明模型是有效的。E,火山图基于CON-FMT组和SD-FMT组相比的差异代谢物筛选。F,火山图基于CON-FMT组和SD+Mel-FMT组相比的差异代谢物筛选。G,火山图基于SD-FMT和SD-FMT组相比的差异代谢物筛选。H,丁酸的相对丰度。I,L-色氨酸的相对丰度。J,热图显示了41种关键代谢物的相对丰度(P < 0.05,VIP > 1)。数据代表均值±SEM,* P < 0.05,** P < 0.01,*** P < 0.001。
4 微生物组组成与表型变量之间的相关性
为了进一步研究代谢物是否被改变,我们用GC/MS测定了粪便中SCFA的含量。结果显示,与CON-FMT组相比,SD-FMT组粪便丁酸含量显著降低(58.9%,P < 0.001)。与SD-FMT组相比,SD+Mel-FMT组粪便丁酸显著增加(146.8%,P < 0.001)。三组间粪便中乙酸和丙酸含量无显著差异(图6B-D)。此外,我们用RT-PCR检测结肠中气单胞菌(Aeromonas)的相对丰度,用ELISA检测海马中LPS的水平。与CON-FMT组相比,SD-FMT组LPS (76.8%,P = 0.017)和气单胞菌(64.0%,P < 0.001)明显增加。与SD-FMT组相比,SD+Mel-FMT组中两者水平明显降低(图6E,F)。相关分析显示,气单胞菌的相对丰度与LPS水平呈正相关(图6G)。粪便丁酸水平与微生物g_Lachnospiraceae_NK4A136_group、g_Eubacterium_xylanophilum、g_lachnospiraceae_a2和s_Clostridiales_bacterium_CIEAF_020呈正相关,与g_Turicimonas和s_Turicimonas_muris呈负相关。这些结果表明,这些微生物的变化可能与粪便丁酸盐水平的变化有关(图6A)。微生物g_Lachnospiraceae_NK4A136_group与在目标区域花费的时间和进入的次数以及IL-10呈正相关,与到达平台的潜伏期和路径长度、TNF-α、IL-6和LPS呈负相关(图6A)。因此,SD或Mel的肠道微生物介导效应可能与其对微生物代谢物(LPS和丁酸盐)潜在的调节作用有关。
图6 微生物组组成与表型变量之间的相关性。
A,顶部关注代谢产物与特征菌群的相关性(Spearman相关检验,FDR < 0.05)。B,粪便中乙酸含量(n = 8)。C,粪便中丙酸含量(n = 8)。D,粪便中丁酸含量(n = 8)。E,海马内LPS水平(n= 7)。F,结肠气单胞菌相对丰度(n = 7)。G,粪便中丁酸盐含量与结肠气单胞菌的相关分析图。x轴为结肠气单胞菌相对丰度;y轴为粪便丁酸盐水平。CON-FMT:接受对照菌群FMT小鼠,SD-FMT:接受睡眠剥夺菌群FMT小鼠,SD+Mel-FMT:接受SD+Mel (20 mg/kg)菌群FMT小鼠。数据代表均值±SEM,* P < 0.05,** P < 0.01,*** P < 0.001。
5 褪黑素可改善A. veronii定植引起的小鼠神经炎症和记忆障碍的发生
为了验证气单胞菌水平升高在SD诱导的记忆障碍中的作用,我们建立了A. veronii定植小鼠模型(图7A)。在A. veronii定植后,我们观察到小鼠到达平台的潜伏期(72.3%,P < 0.001)和路径长度(70.3%,P <0.001)增加,并且在目标区域花费的时间(42.2%,P = 0.001)和进入的次数(57.7%,P = 0.006)减少(图7B-H)。此外,与CON组相比,Aero组海马CA1、CA3和DG区Iba1阳性细胞的IOD分别高23.1% (P = 0.017)、23.9% (P = 0.026)和22.4% (P = 0.023)(图7I,J)。我们还观察到,与CON组相比,Aero组海马中LPS (86.8%,P = 0.002)、IL-6 (69.4%,P < 0.001)和TNF-α (25.0%,P = 0.045)水平显著升高,IL-4 (65.5%,P = 0.002)和IL-10 (54.4%,P = 0.002)水平显著降低(P <0.001)(图7K-O)。与CON组相比,Aero组TLR4 (46.8%,P = 0.005,图7P)、HDAC3 (96.9%,P< 0.001,图7Q)、p-IκB (73.9%,P = 0.005,图7R)、p-P65 (83.5%,P = 0.002,图7S)和cleavedcaspase-3 (54.8%,P = 0.004,图7T)表达水平升高。然而,添加Mel抑制了这一过程,导致CON组和A+Mel组之间没有显著差异。
图7 褪黑素可改善由气单胞菌定植引起的小鼠神经炎症和记忆障碍的发生。
A,实验设计示意图。B,空间记忆测试轨迹图(带隐藏平台)。C,空间记忆测试轨迹图(无隐藏平台)。D,到达平台的潜伏期(n = 8)。E,到达平台的路径长度(n = 8)。F,待在目标区域的时间(n = 8)。G,进入目标区域的次数(n = 8)。H,到达平台的路径效率(n = 8)。I,海马中LPS水平(n = 7)。J-M,海马中细胞因子(TNF-α、IL-6、IL-4和IL-10)的水平(n = 5)。N,不同实验组免疫组化小胶质细胞图像。免疫组化结果用ImageJ处理。比例尺= 50 μm。O,海马CA1、CA3和齿状回(DG)区Iba1阳性细胞的IOD (n = 6)。P–T,海马中TLR4、HDAC3、p-IκB、p-P65、cleaved caspase-3水平(n = 6)。CON:对照组,Aero:气单胞菌定植组,A+Mel:气单胞菌+褪黑素(20 mg/kg)组。数据代表均值±SEM,*P < 0.05,**P < 0.01,***P < 0.001。
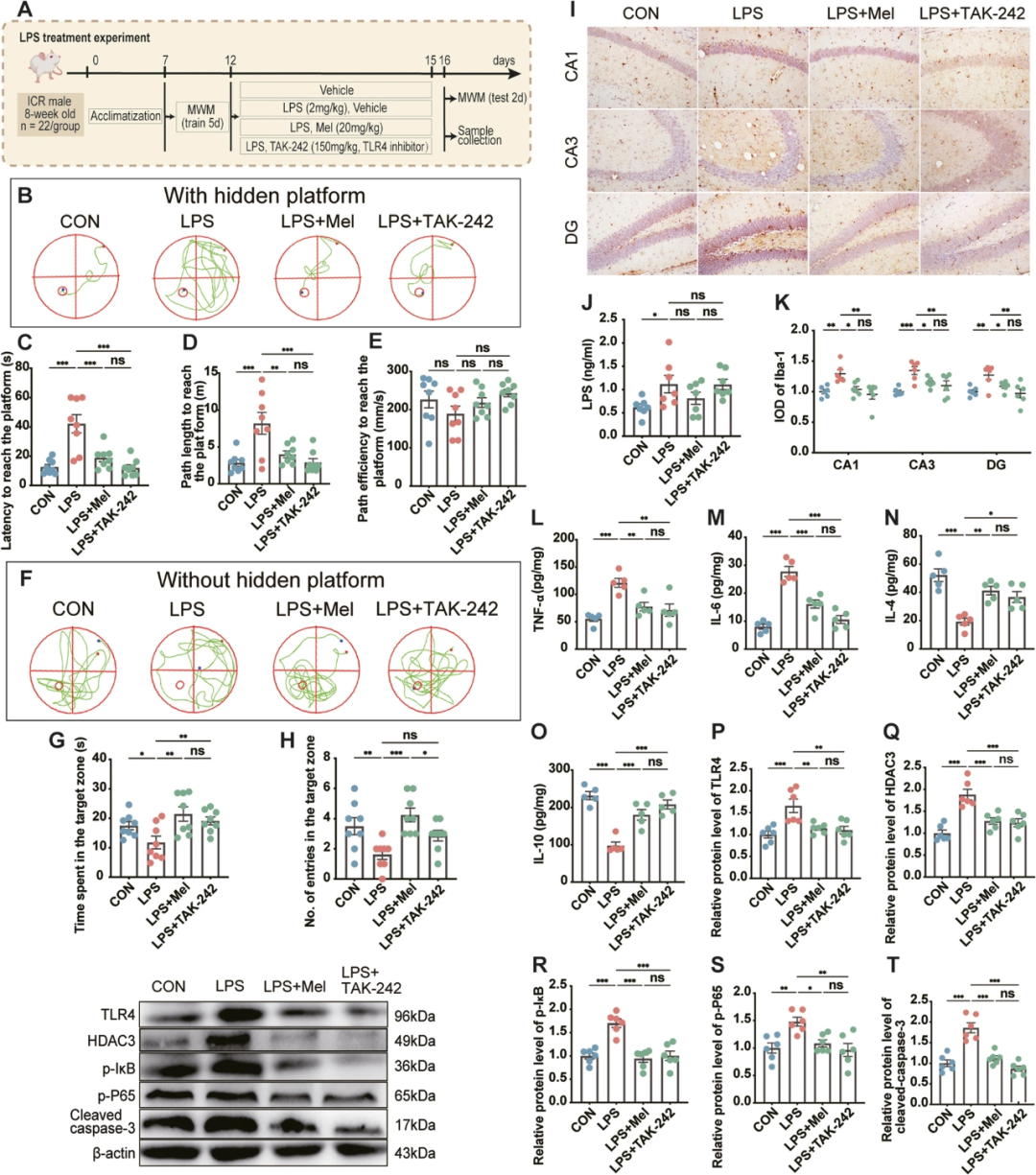
6 褪黑素可改善脂多糖诱导的小鼠神经炎症和记忆障碍
为了评估Mel和LPS介导的记忆损伤的相关性,我们建立了添加或不添加Mel和TAK-242的LPS诱导小鼠模型(图8A)。与CON组相比,LPS处理组小鼠到达平台的潜伏期(230.3%,P < 0.001;图8C)和路径长度(189.1%,P < 0.001;图8D)增加,进入目标区域的时间(32.8%,P = 0.04;图8G)和次数(53.6%,P = 0.005;图8H)减少。此外,LPS (66.6%,P = 0.013;图8J)、TNF-α (54.4%,P = 0.001;图8L)和IL-6 (42.0%,P < 0.001;观察图8M)上调。IL-4 (63.0%,P < 0.001;图8N)和IL-10 (57.8%,P < 0.001;图8O)下调是显而易见的。此外,LPS组海马CA1、CA3和DG区Iba1阳性细胞的IOD分别比CON组高29.5% (P = 0.01)、34.5% (P = 0.001)和27.0% (P = 0.003) (图8I,K)。这些结果表明,与CON组相比,LPS组中TLR4 (65.6%,P = 0.001,图8P)、HDAC3 (88.2%,P < 0.001,图8Q)、p-IκB (70.3%,P < 0.001,图8R)、p-P65 (48.0%,P = 0.007,图8S)和cleaved caspase-3 (86.0%,P < 0.001,图8T)的表达水平增加。添加Mel或TAK-242后,这些变化被逆转,两组之间没有显著差异。
图8 褪黑素可改善小鼠脂多糖诱导的神经炎症和记忆障碍。
A,实验设计示意图。B,空间记忆测试轨迹图(带隐藏平台)。C,到达平台的潜伏期(n = 8)。D,到达平台的路径长度(n = 8)。E,到达平台的路径效率(n = 8)。F,空间记忆测试轨迹图(无隐藏平台)。G,在目标区域花费的时间(n = 8)。H,进入目标区域的次数(n = 8)。I,不同实验组免疫组化小胶质细胞图像。免疫组化结果用ImageJ处理。比例尺= 50 μm。J,海马中脂多糖水平(n = 7)。K,海马CA1、CA3和齿状回(DG)区Iba1阳性细胞的IOD (n = 6)。L-O,海马中细胞因子(TNF-α,IL-6,IL-4和IL-10)的水平(n = 5)。P-T,海马中TLR4、HDAC3、p-IκB、p-P65、cleaved caspase-3水平(n = 6)。CON:对照组,LPS:脂多糖(2 mg/kg)组,LPS+Mel:LPS+褪黑素(20 mg/kg)组,LPS+TAK-242:LPS+TAK-242(TLR4抑制剂,150 mg/kg)组。数据代表均值±SEM,*P < 0.05,**P < 0.01,***P < 0.001。
7 丁酸盐对褪黑素改善SD小鼠记忆障碍的影响
为了研究丁酸盐是否可以介导由Mel引起的SD诱导的认知障碍的改善,我们观察了CON、SD、SD+Abs、SD+Mel、SD+Abs+Mel和SD+Abs+丁酸盐组小鼠在隐藏或可见平台时MWM的变化(图9A)。与CON组相比,SD组和SD+Abs组小鼠的空间记忆功能明显受损。后两组到达平台的路径长度和潜伏期显著增加(P < 0.05;图9B-E),并且在目标区域花费的时间和进入次数显著减少(P < 0.05;图9F-H)。相比之下,补充Mel逆转了SD诱导的空间记忆障碍变化;SD+Mel组、SD+Abs+Mel组和CON组在到达平台的路径长度和潜伏期、花费的时间以及进入目标区域的次数方面均无显著差异(图9B-H)。与补充Mel相似,补充丁酸盐可以改善SD诱导的认知障碍。SD+Abs+丁酸盐组与CON组之间没有观察到任何参数的差异(图9B-H)。此外,各组间路径效率无显著差异(图9E)。由此可见,丁酸盐处理实验结果表明,丁酸盐作为肠道菌群的信号分子,可能介导Mel改善SD引起的认知障碍。
为了研究丁酸盐是否能介导由Mel引起的SD诱导的神经炎症和细胞凋亡的改善,我们检测了海马中Iba1表达的变化以及炎症细胞因子和细胞内信号蛋白的释放。与CON组相比,SD组和SD+Abs组神经炎症明显,海马CA1、CA3、DG中Iba1阳性细胞IOD(P < 0.05;图9I-L)、IL-6、TNF-α水平明显升高,IL-4、IL-10水平显著降低(P < 0.05;图9M-P)。对于细胞内信号蛋白,与CON组相比,SD和SD+Abs组中HDAC3 (P < 0.05;图9Q),p-IκB (P < 0.05;p-P65 (P < 0.05,图9S)和cleaved caspase-3 (P < 0.05,图9T)的表达明显上调。相反,补充Mel逆转了SD引起的神经炎症和细胞凋亡的变化。上述指标在SD+Mel、SD+Abs+Mel、CON组间无明显差异。与补充Mel类似,补充丁酸也能改善SD诱导的神经炎症和细胞凋亡。SD+Abs+丁酸盐组与CON组之间各项参数均无差异。
图9 丁酸盐对Mel改善睡眠剥夺小鼠记忆障碍的作用。
A,实验设计示意图。B,空间记忆测试轨迹图(带隐藏平台)。C,到达平台的潜伏期(n = 8)。D,到达平台的路径长度(n = 8)。E,到达平台的路径效率(n = 8)。F,空间记忆测试轨迹图(无隐藏平台)。G,进入目标区域的次数(n = 8)。H,停留在目标区域的时间(n = 8)。I,不同实验组免疫组化小胶质细胞图像。免疫组化结果用ImageJ处理。比例尺=50 μm。J-L,海马CA1、CA3和齿状回(DG)区Iba1阳性细胞的IOD (n = 6)。M-P,海马中细胞因子(TNF-α、IL-6、IL-4和IL-10)的水平(n = 5)。Q-T,海马中HDAC3、p-IκB、p-P65和cleaved caspase-3的水平(n = 6)。CON:对照组,SD:睡眠剥夺组,SD+Mel:SD+褪黑素(20 mg/kg)组,SD+Abs:SD+抗生素组,SD+Abs+Mel:SD+抗生素+褪黑素组,SD+Abs+丁酸:SD+抗生素+丁酸(40 mM)组。数据代表均值±SEM,* P < 0.05,** P < 0.01,*** P < 0.001。
8 丁酸盐对LPS诱导BV2细胞炎症反应和细胞神经毒性的影响
为了研究LPS或丁酸代谢物在Mel诱导的SD引起的认知障碍中的作用,用LPS处理BV2细胞以模拟神经炎症,并补充丁酸作为干预措施(图10A)。正如预期的那样,BV2细胞暴露于LPS导致TNF-α (283.8%,P < 0.001,图10B)和IL-6 (288.7%,P < 0.001,图10C)的分泌增加,IL-4 (57.8%,P = 0.001,图10D)和IL-10 (35.0%,P < 0.001,图10E)的分泌减少。与对照组相比,LPS处理显著提高了BV2细胞中HDAC3 (68.5%,P = 0.001,图10F)、p-IκB (51.4%,P = 0.001,图10G)和p-P65 (98.7%,P < 0.001,图10H)蛋白的相对表达水平。此外,我们采用小胶质细胞条件培养基(CM)系统来评估丁酸盐减轻小胶质细胞神经毒性是否与神经细胞的存活有关。来自LPS诱导的BV2小胶质细胞的CM,添加或不添加丁酸盐预处理后加入HT22细胞。LPS诱导CM通过上调cleaved caspase-3水平刺激HT22细胞凋亡(42.9%,P = 0.002,图10I)。然而,丁酸盐预处理有效地逆转了这些LPS诱导的变化。相反,与LPS组相比,TAK-242处理后,我们观察到HDAC3 (27.2%,P = 0.001,图10F)、p-IκB(33.2%,P = 0.001,图10G)和p-P65蛋白(36.7%,P < 0.001,图10H)下调。本研究结果表明,与LPS处理组相比,AZD3965抑制MCT1导致LPS + 丁酸盐+ AZD3965处理组中HDAC3 (47.4%,P = 0.001,图10F)、p-IκB (54.3%,P = 0.001,图10G)和p-P65蛋白(92.1%,P < 0.001,图10H)的表达上调。此外,与LPS+丁酸盐组相比,ITSA-1作为HDAC3激动剂处理后,p-IκB (34.8%,P = 0.001,图10G)和p-P65蛋白(88.7%,P < 0.001,图10H)明显上调。对HDAC3蛋白表达水平没有影响。然而,用PDTC(NF-κB拮抗剂)处理可以模仿丁酸盐的有益作用。
图10 丁酸盐对LPS诱导BV2细胞炎症反应和细胞神经毒性的影响。
A,实验设计示意图。B-E,BV2细胞中细胞因子(TNF-α、IL-6、IL-4和IL-10)的水平(n = 5)。F-H,BV2细胞中HDAC3、p-IκB和p-P65的水平(n = 6)。I-H,T22细胞中cleaved caspase-3的相对蛋白水平(n = 6)。TAK-242:TLR4抑制剂,PDTC:NF-κB拮抗剂,AZD3965:MCT1抑制剂,ITSA-1:HDAC3激动剂。数据代表均值±SEM;与对照组相比,* P < 0.05,** P < 0.01,*** P < 0.001。与LPS组相比,#P < 0.05,##P < 0.01,### P < 0.001。与LPS+Mel组相比,+P < 0.05,++P < 0.01, + + +P < 0.001。
讨论
睡眠不足是影响身体多个组织的压力源。实验室研究记录了睡眠不足小鼠的认知障碍和肠道功能障碍,并证明外源性Mel可有效缓解SD诱导的损伤。肠道菌群失调导致大脑功能,如记忆形成和认知功能受损。我们假设,在睡眠不足的小鼠中,肠道和大脑之间存在密切的交流。为了探索这一假设,我们建立了一个FMT模型,以进一步验证肠道微生物在SD引起的记忆障碍中的作用。我们用抗生素处理小鼠,以耗尽大部分肠道菌群,并促进粪便细菌的后续定植。将CON、SD和SD+Mel组小鼠的粪便菌群移植到受体小鼠体内。结果发现被SD菌群定植的受体小鼠到达平台的潜伏期和路径长度增加,进入目标区域的次数和时间减少,这表明SD引起的微生物失调可能会影响认知障碍。与我们的结果类似,接受帕金森病肠道菌群处理的小鼠表现出胃肠功能障碍和运动缺陷。此外,在受体的SN中检测到多巴胺能神经元死亡。同样,我们还发现接受SD菌群的受体小鼠海马中Iba1阳性小胶质细胞数量增加,促炎因子(IL-6和TNF-α)上调,炎症因子(IL-4和IL-10)下调。进一步分析表明,SD-FMT小鼠海马中Bax和cleavedcaspase-3蛋白水平升高,Bcl-2蛋白水平降低。这些结果表明,SD-FMT小鼠海马中存在神经炎症反应和神经元损失,这与SD诱导的脑损伤表型一致。然而,被SD+Mel菌群定植的受体小鼠没有表现出明显的认知障碍,在海马区也没有观察到过度激活的小胶质细胞、神经炎症反应和细胞凋亡。综上所述,Mel可逆转SD诱导的肠道菌群失衡,改善后的肠道菌群不影响正常小鼠的记忆功能。这些结果表明,肠道菌群的变化和由此产生的SD有害症状可以传播。
越来越多的研究证实,肠道微生物群落还可以通过肠-脑轴影响动物的认知功能。进一步的微生物群分析表明,Mel处理的保护作用可能是由正常肠道微生物群的重建介导的。总体而言,当前的α和β多样性结果显示,SD+Mel-FMT组和CON-FMT组的微生物群落相似。此外,CON-FMT组和SD+Mel-FMT组在不同分类单元水平上的比较显示,肠道菌群谱没有显著差异,这表明Mel在SD诱导的小鼠中恢复了健康的菌群。
在FMT实验中,我们的靶向检测显示,相对于CON-FMT组,SD-FMT组的气单胞菌相对丰度显著上调。这一发现与报道的睡眠不足的小鼠中气单胞菌的变化一致。而SD+Mel-FMT组的气单胞菌相对丰度明显低于SD-FMT组。气单胞菌是变形菌门、γ-变形菌纲、气单胞菌纲、气单胞菌科的革兰氏阴性属,是临床上引起肠道和肠外感染的重要人类病原体,其细胞壁成分LPS经研究证明可通过刺激血脑屏障的通透性,越过肠道屏障进入体循环,引起神经炎症。先前的一项研究记录了气单胞菌对小鼠后代发育性运动反射和大脑氧化应激的神经毒性。此外,我们在SD-FMT小鼠中观察到Proteobacteria和Gammaproteobacteria显著上调。因此,我们推测气单胞菌可能介导急性SD诱导的小鼠认知障碍。在本研究中,被气单胞菌定植的小鼠表现出受损的SD样认知功能,并显著增加海马中的LPS水平。进一步的测试显示,海马中的神经炎症反应和细胞凋亡增加,如小胶质细胞激活、促炎细胞因子增加、抗炎细胞因子减少,以及cleaved caspase-3水平的增加,Mel的补充改善了这些指标的变化。综合研究结果表明Mel可以改善由气单胞菌定植引起的记忆障碍。
上述结果表明气单胞菌可介导SD诱导的小鼠认知功能障碍。我们还观察到SD-FMT小鼠海马中LPS水平升高。相关分析显示,结肠气单胞菌相对丰度与海马中LPS水平呈正相关。据报道,在有噬菌体的情况下,LPS破坏肠屏障的稳定性并进入体循环。一方面通过血液循环到达身体多个组织,另一方面可以激活免疫细胞释放大量炎症因子,加重全身炎症的发生。循环的LPS和释放的炎症因子使血脑屏障不稳定并增加其渗透性。然后LPS可以破坏血脑屏障并进入脑实质。因此,我们假设LPS介导SD诱导的小鼠认知功能障碍。为了支持这一假设,用LPS处理的正常小鼠表现出SD样认知障碍,在海马中观察到LPS含量显著增加,伴随着小胶质细胞的过度激活和促炎因子的大量释放。与我们的结果相似,先前也在接受睡眠剥夺者的FMT的小鼠中检测到高水平的LPS。进一步的机制研究发现,TLR4、p-P65、p-IκB和cleaved caspase-3在气单胞菌定植小鼠和LPS处理小鼠海马细胞中的表达水平显著升高。外源添加Mel可有效逆转这些变化。这些结果表明Mel可以减轻气单胞菌和LPS引起的海马神经炎症,最终改善SD样认知障碍。
本研究观察到Mel给药后SD诱导小鼠的健康菌群恢复;由此产生的微生物群与对照组相似。有趣的是,我们观察到在移植了SD+Mel微生物的受体小鼠中,产生丁酸盐的细菌大量增加,包括Lachnospiraceae_NK4A136_group、Eubacteriumxylanophilum、Ruminococcus_1和Lachnospiraceae_A2。此前有研究发现,Lachnospiraceae_NK4A136_group的丰度可以改善衰老大鼠的肠道屏障功能。该组是主要产生丁酸盐的细菌之一,其丰度与炎症水平呈显著负相关。此外,真杆菌种类,如E. rectale和E. eligens,已与较低的脆弱性,提高认知能力,以及增加SCFA和支链脂肪酸的生产等几个标记物呈正相关。真菌与炎症标志物IL-2、c反应蛋白呈负相关。Ruminococcus_1也与抑郁样行为的减少呈正相关。我们还通过GC-MS发现,不同组间的代谢物变化明显,与微生物的变化相似。本研究中,与SD-FMT组相比,SD+Mel-FMT组显著增加574种代谢物,减少26种代谢物。其中,丁酸和L-色氨酸是SD+Mel-FMT组显著上调的代谢物。进一步的LC-MS分析表明,SD-FMT小鼠粪便中丁酸含量显著降低,而乙酸和丙酸含量未见变化。然而,移植SD+Mel小鼠的微生物群显著恢复了SD诱导的丁酸盐减少。这一发结果表明,增加微生物丁酸盐的产生可能在调节Mel对SD引起的认知障碍的肠道微生物相关效应中起主导作用。
重要的是,丁酸盐作为抗炎活性的关键介质,在SCFAs中受到了最广泛的关注。最近的一项研究表明,口服丁酸盐可改善阿尔茨海默氏症小鼠的神经炎症和认知障碍。在当前的研究中,丁酸盐或Mel被给予睡眠不足的小鼠,以进一步验证丁酸盐的有益作用。结果表明,补充丁酸盐或Mel可以有效地减轻睡眠不足小鼠海马中Iba1阳性细胞的数量。促炎细胞因子和促凋亡蛋白水平显著增加,最终逆转小鼠的认知功能。但我们前期研究表明,SD小鼠补充Mel (SD+Mel和SD+Abs+Mel组)后,Mel和丁酸含量显著升高,而补充丁酸(SD+Abs+丁酸组)仅恢复了丁酸含量,并没有消除SD对Mel分泌的抑制。这些观察结果表明丁酸盐作为肠-脑轴的信号分子,可以介导Mel对SD诱导的记忆障碍的改善作用。
小胶质细胞是大脑中的先天免疫细胞,在调节大脑免疫反应中起着重要的作用。进入大脑的LPS与小胶质细胞上的TLR4受体结合,通过MyD88启动下游IκB/NF-κB或丝裂原活化蛋白激酶/细胞外信号活化激酶(MAPK/ERK)信号转导通路,激活相关蛋白,促进促炎因子TNF-α和IL-6的表达。大量的分泌产生强烈的炎症作用。在大脑中,丁酸盐可以通过转运蛋白通过细胞膜进入细胞,并通过抑制组蛋白去乙酰化酶发挥抗炎作用。相反,丁酸盐通过与受体结合并激活下游信号通路产生抗炎作用。MCT1转运体在小胶质细胞中广泛表达,而G蛋白偶联受体在小胶质细胞中表达较低。因此,我们推测丁酸盐和LPS这两种与炎症密切相关的代谢产物可能以小胶质细胞为靶细胞来调节大脑中的炎症反应。与对照组相比,LPS处理的BV2细胞培养液中促炎细胞因子分泌增加,抗炎细胞因子分泌减少,HDAC3、p-IκB和p-P65含量增加。在HT22细胞中,cleaved caspase-3蛋白的增加提示细胞凋亡的发生。然而,丁酸盐处理逆转了脂多糖诱导的变化。此外,TLR4抑制剂TAK-242和NF-κB拮抗剂PDTC的加入模拟了丁酸盐对LPS诱导的BV2细胞炎症反应的改善作用。补充MCT1抑制剂AZD3965和HDAC3激动剂ITSA-1阻断了丁酸盐的保护作用。
结论
本研究的数据揭示了Mel对SD诱导的认知障碍的保护作用。进一步的机制研究表明,下调Aeromonas和LPS水平,上调LachnospiraceaeNK4A136和丁酸盐水平可能是Mel在SD引起的认知障碍中发挥神经保护作用的潜在机制(图11)。
图11 褪黑素通过微生物-肠-脑轴对睡眠剥夺引起的认知障碍的保护作用示意图。
简单地说,肠道微生物及其代谢产物介导褪黑素对SD诱导的认知障碍的改善作用。一种可行的机制是Mel通过重塑肠道菌群稳态,下调了气单胞菌的数量和组成成分LPS的产量,并上调了Lachnospiraceae_NK4A136的数量和丁酸代谢物的产量。这些事件抑制TLR4/HDAC3/NF-κB信号通路,从而预防神经炎症,最终减轻睡眠剥夺小鼠的神经元凋亡和记忆障碍。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
(京)网药械信息备字(2025)第00082号
京公网安备 11010802020745号
工商备案公示信息
(京)网药械信息备字(2025)第00082号

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您









































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




