
生物信息学和免疫信息学工具预测MP3RT是一种无毒、无致敏性的肽基疫苗,具有较高的抗原性和免疫原性。
作者:程鹏,薛勇,王杰,贾再兴,王亮,龚文平
第一作者及单位:程鹏,结核病防治重点实验室/结核病诊疗新技术北京市重点实验室,中国人民解放军总医院第八医学中心结核病医学部,北京
通讯作者及单位:龚文平,王亮:结核病防治重点实验室/结核病诊疗新技术北京市重点实验室,中国人民解放军总医院第八医学中心结核病医学部,北京
Evaluation of the consistence between the results of immunoinformatics predictions and real-world animal experiments of a new tuberculosis vaccine MP3RT
Cheng P,Xue Y,Wang J,Jia Z,Wang L, Gong W.
Front Cell Infect Microbiol,2022,12:1047306.
doi: 10.3389/fcimb.2022.1047306. eCollection 2022.
PMID: 36405961
背 景
结核病是一种古老的传染病,一直威胁着人类的生命和健康。然而,到目前为止仍然没有更好的办法对结核病进行控制和预防。接种疫苗是预防和控制结核病和其他分枝杆菌感染的有效方法。卡介苗 (BCG)发明于19世纪,是唯一获得许可的结核病疫苗。虽然卡介苗对儿童重度结核和粟粒结核具有良好的保护作用,但其有效保护期只有10~20年。因此,迫切需要开发新的结核病疫苗,以弥补卡介苗的不足。在先前的研究中我们开发了一种新的肽基疫苗MP3RT,并在人源化和野生型小鼠上评估了其免疫特异性和保护效率。与上述传统结核疫苗相比,肽基疫苗具有抗原性好、主要组织相容性复合体(MHC)分子识别效率高、副作用少、生产简单、运输方便等显著优势。生物信息学和免疫信息学技术的快速发展为反向疫苗学的兴起提供了机会。免疫信息学的迅速发展促进了当代免疫学和新疫苗研究的进步。它被广泛应用于表位发现、精确免疫工程和加速疫苗设计。反向疫苗学是基于免疫信息学和分子生物学,从全基因水平筛选具有保护性免疫应答的候选抗原的方法。该策略具有方便、范围广、安全、省力、节约成本等优点。然而,免疫信息学预测与MP3RT疫苗的真实动物实验结果之间的一致性仍不清楚。因此,本研究利用多种生物信息学和免疫信息学工具预测MP3RT疫苗的抗原性、免疫原性、二级结构和理化性质。此外,我们还分析了其功能预测,如三级结构建模、分子对接和免疫刺激。最后,我们通过动物实验验证了MP3RT疫苗的免疫学特性,并通过对比分析现实数据和生物信息学预测数据,评估了生物信息学技术在反向疫苗研究中的价值。
研究方法
一、生物信息学预测
1. MP3RT分子序列
在我们之前的研究中,MP3RT疫苗由6个免疫优势肽(Mtb8.469-83、PPE18115-129、PPE18149-163、PPE68138-152、RpfA377-391和TB10.421-35)组成,许多研究已经确定这五种抗原是潜在的结核病疫苗候选抗原。

2. MP3RT疫苗的免疫原性、抗原性、致敏性和毒性预测
疫苗的免疫原性和抗原性在抗击结核分枝杆菌感染方面起着至关重要的作用。因此,使用免疫表位数据库(IEDB)免疫原性服务器(http://tools.iedb.org/immunogenicity/)预测MP3RT疫苗的免疫原性。VaxiJen v2.0 (http://www.ddg-pharmfac.net/vaxijen/VaxiJen/VaxiJen.html)对MP3RT疫苗的抗原性进行预测,并使用ANTIGENpro服务器(http://scratch.proteomics.ics.uci.edu/)对其进行进一步验证。
为了更好地验证MP3RT疫苗的毒副作用,使用AllerTOP v.2.0服务器(http://www.ddg-pharmfac.net/AllerTOP/)和Allergen FP v.1.0服务器(http://ddg-pharmfac.net/AllergenFP/)预测MP3RT疫苗的敏感性。最后,使用Toxin Pred服务器(http://crdd.osdd.net/raghava/toxinpred/)预测MP3RT疫苗的毒性。AllerTOP v.2.0服务器和Toxin Pred服务器的预测结果分别显示为“致敏性”或“非致敏性”和“无毒性”或“有毒性”。
3. MP3RT疫苗的理化性质和溶解度
使用ExPASy ProtParam服务器(https://web.expasy.org/protparam/)预测MP3RT疫苗的理化参数,包括理论等电点(pI)、体内外半衰期、不稳定性指数等理化性质。 然后,使用Protein-Sol服务器(https://protein-sol.manchester.ac.uk/)预测MP3RT疫苗的溶解度。
4. 二级结构和三级(3D)结构预测和优化
使用PSIPRED服务器(http://bioinf.cs.ucl.ac.uk/psipred/)预测了MP3RT疫苗的二级结构。接下来,使用3Dpro服务器[Scratch Protein Predictor (uci.edu)]预测MP3RT疫苗的3D空间排列。随后,通过GalaxyRefine web服务器(https://galaxy.seoklab.org/cgi-bin/submit.cgi?type=REFINE)对初始模型进行优化,提高了全局和局部结构质量。
5. 三级结构验证与分子对接
使用ERRAT服务器(https://saves.mbi.ucla.edu/)评估MP3RT疫苗3D结构的不确定性。此外,通过SWISS-MODEL服务器(https://swissmodel.expasy.org/assess)绘制Ramachandran图。
为了评估MP3RT疫苗与HLA-DRB1*01:01等位基因、TLR-2和TLR-4受体之间的亲和力,使用ClusPro2.0服务器(https://cluspro.bu.edu/home.php)进行配体-受体对接分析。最后使用PyMOL2.5.3软件(Schrödinger, New York, USA)进行可视化分析。
6. 构象B细胞表位的预测
构象B细胞表位是由蛋白质折叠产生的,蛋白质折叠使远处的残基彼此靠近。通过ElliPro服务器(http://tools.iedb.org/ellipro/)预测了MP3RT疫苗的构象B细胞表位。
7. 免疫模拟
使用C-ImmSim服务器(https://150.146.2.1/C-IMMSIM/index.php)进行免疫刺激,该服务器可以预测MP3RT疫苗刺激后T和B淋巴细胞及细胞因子的表达变化。
二、实验验证
1. 实验动物
本研究所使用的雌性人源化C57BL/6小鼠(HLA-A11+/+DR1+/+H-2-b2m-/-/IAb-/-)由北京微生物与流行病学研究所(北京,中国)周育森教授赠送。所有动物实验遵循中国国家科学技术委员会制定的《实验动物管理条例》,所有小鼠的处死都在麻醉状态下进行。本研究由解放军总医院第八医学中心动物伦理委员会批准(批准文号:309201808171015)。
2. 小鼠免疫
人源化小鼠被随机分为两组:对照组和MP3RT组。对照组(n = 6)小鼠接受30 μg CpG寡核苷酸(ODN2395)佐剂(100 μl PBS)。MP3RT组(n = 7)小鼠接受30 μg CpG寡核苷酸(ODN2395)佐剂+30 μg MP3RT CpG佐剂(100 μl PBS)。初次免疫后,MP3RT组小鼠于第28天和第42天均接种20 μg MP3RT+20 μg CpG(100 μl PBS),对照组小鼠分别于第28天和第42天接种20 μg GpG佐剂(100 μl PBS)。分别于第一次免疫后第0、14、28、42、56、70天采集小鼠血液。最后,在首次疫苗接种后第84天处死小鼠,摘取小鼠脾脏。
3. MP3RT特异性抗体检测
每只小鼠血样2500转离心10分钟,分离血清-80℃保存。为了减少个体差异,将各组小鼠的血清汇集起来,分别进行四份抗体检测。按照我们之前的研究,使用间接酶联免疫吸附试验(ELISA)检测MP3RT疫苗的特异性抗体IgG及其亚型IgG1和IgG2a。
4. 酶联免疫斑点(ELISPOT)试验
MP3RT组和PBS组小鼠在初始免疫84天后处死。根据我们前期的研究,收集每只小鼠的脾脏,制备10 ml脾脏细胞悬液。然后,按照制造商的说明,使用小鼠IFN-γELISOPTPLUS (Mabtech AB, Nacka Strand, Sweden)测定γ-干扰素(IFN-γ)+T淋巴细胞的频率。最后,斑点形成细胞(SFCs)用CTL-s5 Versa ELISPOT Reader (CTL, Cleveland, OH, USA)进行确认。为减少个体差异,将各组小鼠脾细胞收集在一起,一式四份进行IFN-γ+ T淋巴细胞检测。
5. 细胞因子检测
采用上述方法制备小鼠脾细胞。然后,按照制造商的说明,使用小鼠Th1/Th2/Th17细胞因子试剂盒(BD Biosciences, San Jose, CA, USA)检测白细胞介素-2 (IL-2)、IL-4、IL-6、IL-10、IFN-γ、肿瘤坏死因子-α (TNF-α)和IL-17A等细胞因子。为减少个体差异,将各组小鼠脾细胞聚集在一起,一式四份进行细胞因子检测。
6. 统计分析
使用GraphPad Prism 9.4.0 (San Diego, CA, USA)分析MP3RT特异性抗体、SI和细胞因子的数据。总之,MP3RT疫苗诱导的细胞因子水平按正态性采用Unpaired t检验或Mann-Whitney U检验进行分析。数据以均数±标准误差(SEM)表示,P值< 0.05为差异有统计学意义。
结 果
1. MP3RT疫苗的免疫原性、抗原性、致敏性和毒性
IEDB免疫原性服务器和VaxiJen v2.0服务器预测的MP3RT疫苗的免疫原性和抗原性指数分别为0.60875和0.8309。随后,在ANTIGENpro服务器上进一步验证MP3RT疫苗的抗原性结果为0.875498。此外,AllerTOP v. 2.0服务器、Allergen FP v. 1.0服务器和Toxin Pred服务器的预测结果表明,MP3RT疫苗是无致敏和无毒的。这些数据表明,MP3RT是一种具有高抗原性、强免疫原性、无毒、无致敏作用的潜在结核疫苗。
2. MP3RT疫苗的理化性质和溶解度
疫苗的理化性质显著影响了免疫功能的发展。我们通过ExPASy ProtParam服务器获得了MP3RT疫苗的理化参数,结果如表2所示。通过Protein-Sol server预测MP3RT疫苗的溶解度为0.548(图1A),表明该疫苗具有良好的溶解度。这一预期结果与我们之前研究的实验结果一致。

3. MP3RT疫苗的二级结构、三级结构和三级结构优化
根据PSIPRED服务器,我们预测了MP3RT疫苗的二级结构(图1B),结果表明该疫苗由53.36%的α-螺旋,10.6%的β-链和36.04%的随机线圈组成。MP3RT疫苗模型的3D结构最初使用3Dpro服务器进行设计(图1C)。随后,使用Galaxy WEB服务器对MP3RT疫苗模型进行优化。根据GDT-HA和MolProbity值对优化模型进行排序(表3),GDT-HA值越高,MolProbity值越低表示模型质量越好。MolProbity由3个部分组成:每1000个原子中的原子碰撞数、旋转分子异常值的百分比和Ramachandran偏爱的骨架扭转角。最后,我们选择模型5(图1D)进行进一步研究。

4. 三级结构验证和分子对接
使用ERRAT服务器对MP3RT疫苗的三维结构模型进行验证。MP3RT疫苗的总体质量因子由64.8352%提高到81.746%。Ramachandran图分析发现,完全允许区域从91.34%上升到98.22%,最大允许区域从3.15%下降到0.36%,不允许区域从5.32%下降到0.99%(图2A和图2B)。使用ClusPro2.0服务器进行配体-受体对接分析。结果表明,MP3RT疫苗对HLA-DRB1*01:01、TLR-2和TLR-4受体的最低结合能分别为-1234.1 kcal/mol、-1066.4 kcal/mol和-1250.4 kcal/mol。MP3RT疫苗与HLA-DRB1*01:01、TLR-2或TLR-4受体之间的配体-受体对接的可视化如图3所示。


5. 构象B细胞表位
ElliPro服务器的结果发现共形成了4个构象B细胞表位,其评分分别为0.817(图4A)、0.729(图4B)、0.712(图4C)、0.668(图4D)。

图4
6. MP3RT疫苗对固有免疫细胞的免疫模拟刺激
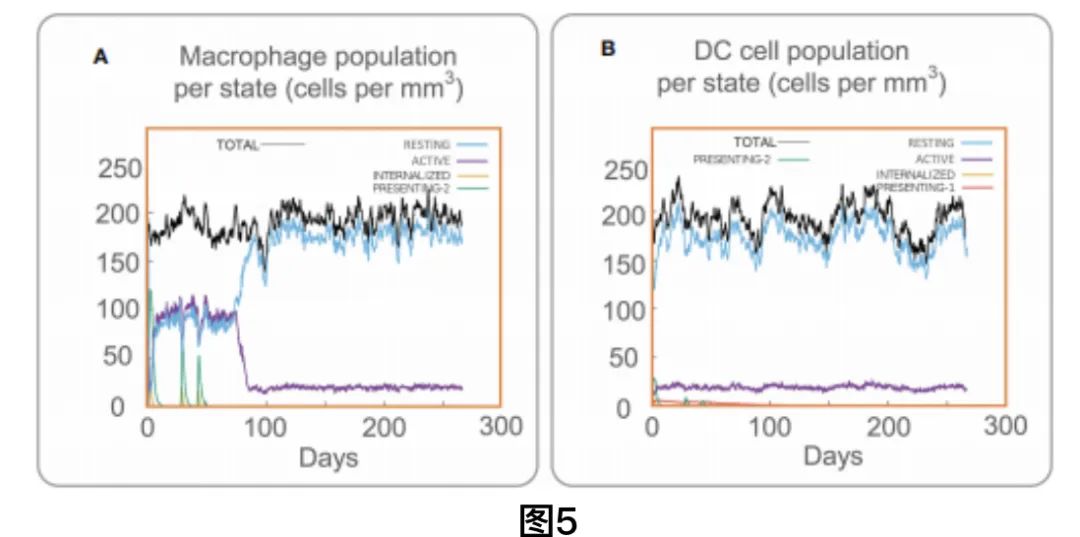
C-ImmSim服务器结果显示MP3RT疫苗可以显著激活巨噬细胞(图5A)。结果表明,从第一次注射到第三次注射,活性巨噬细胞数量维持在100 cells/mm3。第一次接种疫苗80天后,活跃巨噬细胞数量迅速减少,并维持在20 cells/mm3。此外,present-2巨噬细胞数量形成了3次高峰,分别是第2天的120 cells/mm3,第28天的70 cells/mm3,第44天的50 cells/mm3。树突状细胞(dc)是最强的抗原呈递细胞(APCs),在向T细胞呈递方面发挥着重要作用。因此,我们发现在MP3RT疫苗刺激后,总树突状细胞的数量维持在200 cells/mm3,而活跃树突状细胞的数量迅速分泌为20 cells/mm3。与巨噬细胞一样,present-2 DC细胞在3次MP3RT刺激后也形成3个高峰(图5B)。
7. MP3RT疫苗对适应性免疫细胞的免疫模拟刺激
适应性免疫细胞在抵抗结核病发病机制中起着重要作用,主要包括CD4+ T细胞、CD8+ T细胞和B细胞。我们的结果表明,在MP3RT疫苗刺激后,总TH细胞、非记忆TH细胞和记忆TH细胞形成了3个越来越高的峰值,最后在第三次刺激后达到了最高峰(图6A,分别为12000 cells/mm3、10000 cells/mm3和11000 cells/mm3)。与此同时,在第三次刺激后,活跃和静止CD4+ T细胞的数量分别达到8500 cells/mm3和4000个/mm3的峰值(图6B)。有趣的是,复制状态的细胞也形成了3个峰值,在第二次刺激后达到了2200 cells/mm3的最高峰(图6B)。
细胞毒性T淋巴细胞可通过产生穿孔素、颗粒酶B和其他细胞毒性因子清除结核分枝杆菌。C-ImmSim服务器的结果显示,非记忆细胞毒性T淋巴细胞在3次刺激后形成了1150 cells/mm3的峰值(图6C)。然而,当活跃的T淋巴细胞的数量在初级免疫后第50天达到峰值(900 cells /mm3)时,复制的细胞毒性T淋巴细胞的数量达到谷底(200 cells /mm3) (图6D)。我们还预测了MP3RT疫苗接种后B细胞的数量。结果显示,在第三次刺激后,总B细胞的每个状态的数量达到峰值690个/mm3(图6E)。此外,活性B细胞的数量在第三次刺激后达到峰值(690 cells /mm3),present-2细胞的数量在第一次刺激后达到峰值(650 cells /mm3)(图6F)。

8. 比较真实实验与计算机分析的MP3RT疫苗诱导的体液免疫应答
在计算机分析中发现,MP3RT免疫产生了明显更高水平的MP3RT特异性抗体。IgG1+IgG2抗体水平在第三次免疫后达到峰值(650000/ml),然后逐渐下降(图7)。此外,IgG1和IgG2抗体水平分别为250000/ml和23万/ml(图7)。在体内分析中,我们进行了动物实验,将生物信息学预测的免疫谱与MP3RT疫苗的真实实验数据进行比较。分别在第0天、第28天和第42天接种疫苗(图8A)。结果显示,MP3RT免疫小鼠的MP3RT特异性IgG(图8B)、IgG1(图8C)和IgG2a(图8D)水平明显高于PBS免疫小鼠。我们还发现,与第一次免疫后相比,IgG2a/IgG1的比值更高(图8E)。

9. 比较真实实验与计算机分析的MP3RT疫苗诱导的细胞因子反应
计算机分析结果表明,MP3RT接种可显著提高IFN-γ水平。IFN-γ水平在第二次免疫时达到最高值(470000 ng/ml)。与免疫细胞群体不同的是,IFN-γ、IL-2、TGF-β、IL-10、IL-12等细胞因子水平在第二次免疫刺激后达到峰值(图9)。在体内分析,ELISPOT结果显示,MP3RT免疫小鼠的IFN-γ+ T淋巴细胞频率明显高于PBS免疫小鼠(图10A),MP3RT组SI值明显高于PBS组(图10B,P=0.0286)。此外,我们还探索了MP3RT疫苗诱导的细胞因子,如IFN-γ(图11A)、TNF-α(图11B)、IL-2(图11C)、IL-4(图11D)、IL-6(图11E)、IL-10(图11F)和IL-17A(图11G)。我们发现,MP3RT疫苗接种小鼠的IFN-γ(P=0.0286)、IL-2 (P=0.0286)和IL-6 (P=0.0233)水平显著高于PBS疫苗接种小鼠。

研究意义
生物信息学和免疫信息学工具预测MP3RT是一种无毒、无致敏性的肽基疫苗,具有较高的抗原性和免疫原性。我们的研究表明,生物信息学和免疫信息学工具在预测肽基疫苗的理化性质、分子结构和免疫特性方面表现出了良好的性能,并且其预测的体液和细胞免疫反应的结果与真实的动物实验数据高度一致,表明生物信息学和免疫信息学技术可以在反向疫苗学领域推广。
注:除非特别声明,本公众号刊登的所有文章不代表《中国防痨杂志》期刊社的观点。
编辑:王 然
审校:郭 萌
发布日期:2022-12-06
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您
































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




