
尽管对肝癌起源和详细分子特征的了解取得了长足进步,但能够为患者带来实质性生存和改善生活质量的治疗选择仍然很少。
肝脏是原发性肿瘤第六大好发部位,其多由肝硬化或炎症引起。同时,肝脏亦是其他肿瘤(尤其是大肠肿瘤)的常见转移器官,这既是因为肝脏解剖位置及结构特殊,也是因为其具有独特的代谢及免疫抑制环境。在慢性炎症或代谢条件改变等病理状态下,肝脏微环境会发生适应性改变。这样的改变可能对肝脏原发性肿瘤的发生发展产生正反两方面影响,同时也对肿瘤向肝脏或在肝脏内的转移产生作用。因此,2021年7月Xin Li及 Pierluigi Ramadori等人在《Nature Reviews: Cancer》杂志上发表了一篇题为《The immunological and metabolic landscape in primary and metastatic liver cancer》的综述,概述了肝脏生理、免疫及其代谢特性,并于其中特别关注了特定病因对肝脏稳态的影响,以及这样的微环境改变对肿瘤发生、发展及转移的影响。现介绍如下:
癌前肝脏微环境
原发性肝癌由慢性炎症引起,其触发和维持的因素有:免疫细胞激活、固有免疫细胞生成促炎细胞因子、内在肝细胞类型(包括肝细胞和肝窦内皮细胞(LSECs))的稳定性被破坏、脂肪酸的脂毒性和过量铁沉积。上述因素导致ROS和活性氮过度产生,破坏相关机制稳定性,导致DNA损伤修复功能障碍。持续的再生反应导致肝星状细胞(HSCs)激活、胶原蛋白沉积、纤维化进而导致肝硬化。
肝脏的免疫抑制环境
肝脏具有免疫抑制特性,这会削弱有效的T细胞介导的抗原反应,并由常驻肝细胞群维持,包括枯否细胞(KCs)、HSCs、树突状细胞(DCs)、Treg细胞和LSECs。
KCs表达PDL1和低水平的共刺激分子,并可启动Treg细胞。细菌等使KCs抗炎极化,进一步降低LSECs上MHC的表达,限制其免疫刺激能力。
HSCs通过产生抗炎细胞因子TGFβ促进肝脏的免疫耐受。可激活组织再生途径并使炎性单核细胞向髓源性抑制细胞(MDSCs)分化。
肝DCs激活T细胞的效果较其他组织DCs差。肝脏有两种主要的DCs亚型:类浆细胞衍生的DCs似乎介导肝脏免疫耐受,但CD141+常规DCs可归于促炎免疫表型。
肝脏驻留自然杀伤 (NK) 细胞在感染和肿瘤免疫监视中具有异质性。肝脏NK细胞主要分2个细胞群:常规和肝脏特异性驻留NK细胞。肝脏特异性驻留NK细胞通过IFNγ产生细胞毒性,但它们会因肝脏免疫抑制环境保持在低反应状态。MDSCs可产生免疫抑制细胞因子并表达精氨酸酶,抑制T细胞增殖、ROS产生和NK细胞功能。
Treg细胞表达高水平的CD25和CTLA4,与潜在的效应T细胞竞争IL-2、CD80和CD86。它还产生抗炎细胞因子以维持免疫抑制特性。
最后,LSECs表达PDL1、模式识别受体和粘附分子(如ICAM1和VCAM1),介导淋巴细胞向组织浸润和MHC I类或II类表达。但在肝脏免疫抑制状态下,T细胞无应答。
肝脏代谢微环境
肝脏由肝小叶组成。其内肝细胞呈同心圆式分布。血流向心性流过肝窦,使门管区向中央静脉形成递减的氧分压梯度。肝细胞在小叶中空间分布不同,其微环境、功能、酶分布也不同。肝细胞基因表达在时间上也受到调节。
肝细胞功能随空间组织。酒精性肝损伤主要局限在近中心区域。而最常见的非酒精性脂肪性肝病(NAFLD)中,游离脂肪酸及其代谢物引起脂毒性,在近中心区域导致肝细胞骨架受损,失去维持正常细胞周期或程序性死亡的能力。反复的细胞死亡-增殖周期会诱导氧化应激和KCs激活。慢性炎症也会改变肝脏代谢。
细胞死亡作为坏死性炎症的驱动因素
肝细胞死亡是伴随着慢性肝病的过程。它主要以细胞凋亡、坏死性凋亡或坏死的形式出现。尚有其他细胞死亡途径参与,如铁死亡、氧死亡和焦亡。细胞死亡的类型会影响发展成原发性肝癌的类型。
细胞外刺激通过激活细胞死亡受体来触发外源性细胞死亡。细胞内的毒性条件,如高浓度ROS、DNA损伤或由于再生能力耗竭导致的复制压力,会激活与外源性类似的内源性细胞凋亡。
细胞死亡可分为受控(坏死性凋亡)和不受控(坏死)亚型。坏死性凋亡被外在因素激活,也可由内在DNA感受器激活。它诱导RIPK3激活,磷酸化MLKL,形成孔状多聚体,整合到质膜导致质膜破坏。而坏死是在诸如物理、化学或热诱导损伤等应激条件下出现的,它直接破坏细胞完整性。
不同病因刺激会引发不同的细胞死亡途径。慢性炎症诱导的程序性细胞死亡和DNA损伤修复,与基因组稳定性相关分子之间存在直接联系。caspase 8参与慢性炎症诱导的细胞死亡,同时又参与DNA损伤修复。caspase 8复合物触发γH2AX磷酸化,维持DNA完整。获得抗凋亡特性的肝细胞会下调caspase 8导致DNA损伤累积。
死亡肝细胞释放大量损伤相关分子,加上过量酒精或脂肪酸分解代谢产物,这些“内源性抗原”导致免疫耐受的平衡被破坏。KCs打开模式识别受体信号,产生多种促炎细胞因子以及包括CCL1、CCL2和CCL5在内的趋化因子。这导致ICAM1和VCAM1表达增加,以及LSEC上PECAM1的下调,将单核细胞、中性粒细胞和血小板募集到应激部位。在驻留的天然免疫细胞激活后,适应性免疫细胞和炎性单核细胞被募集到炎症肝脏中。
肝脏的肿瘤转变
肿瘤的发生是正常细胞获得生存优势并逐渐积累致癌突变的过程。在有利于基因突变积累和存在诱导致癌信号通路的环境中,肝细胞代偿性再生的增殖细胞更易被赋予恶性潜能。
促肿瘤炎症信号
持续炎症状态下,由KCs分泌的TNF激活JNK和WNT-β-连环蛋白信号通路,促进HCC发生。肝癌中发现淋巴毒素激活了NF-κB通路。其他炎症介质包括IL-1β、IL-6、IL-11和IL-23也是促肿瘤细胞因子。此外,炎症环境中Ang2、VEGF、CXCL1和CXCL8的富集可以刺激血管生成。
持续代偿性细胞死亡和增殖会加剧氧化应激,导致DNA修复机制受损。ROS或活性氮抑制DNA修复中的关键酶。肝癌中最常见的TP53突变由氧化应激诱导,它通过表观遗传,诱导错配修复酶和肿瘤抑制因子失活。氧化应激又是炎症的结果。HCC中,活化的免疫细胞增加ROS等产生,激活STAT1和STAT3信号通路。STAT1上调CXCL9和脂质运载蛋白2,促进肝脏炎症;STAT3似乎通过IL-6–STAT3自分泌环,驱动癌症祖细胞的恶性转化。
肝细胞的可塑性及转化
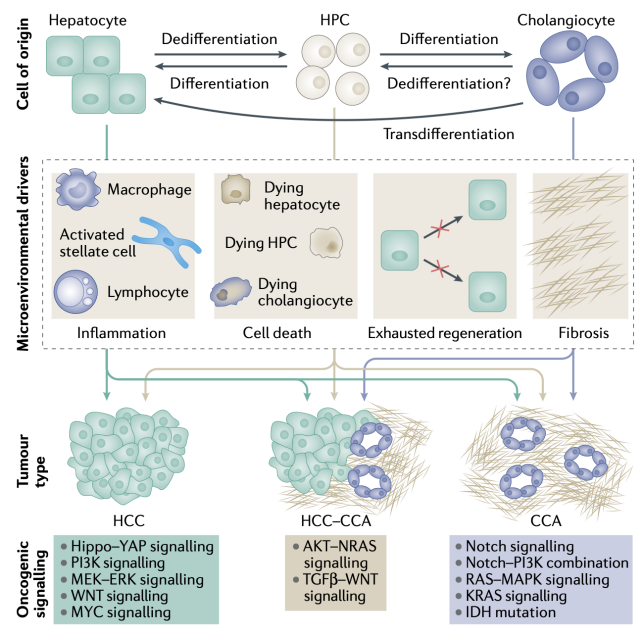
混合HCC和CCA表型的肿瘤正在逐渐增加,其恶性程度升高且预后不良。最近发现肝脏肿瘤发生中的谱系分化其实具有可塑性。
微环境可以影响不同细胞类型的谱系决定。Jagged1-Notch信号通路是肝细胞可塑性的主要调节因子。另一个是WNT-β-连环蛋白信号通路。胆管细胞中WNT-β-连环蛋白信号促进肝细胞通过旁分泌信号转分化为胆管细胞。肝细胞线粒体功能障碍会导致ROS产生,激活巨噬细胞释放TNF并导致胆管细胞中的JUN-JNK通路激活,从而导致CCA发展。分子分析表明,多数CCA显示出胆管细胞特异性模式,但其余则具有肝干细胞的特征,这表明肝干细胞也能够产生CCA。微环境中的特定细胞因子谱可以分别驱动转化的肝细胞进入HCC或CCA谱系。
肝癌的代谢情况
在肝肿瘤中经常观察到代谢的改变。肿瘤细胞倾向糖酵解来满足能量和合成代谢需求,而所有代谢产物的代谢途径在肝癌中都会失调。
代谢重编机制肝癌中至关重要,代谢底物的改变一方面促进了肝细胞增殖,另一方面改变了免疫活性。这不仅使肝癌在实质内增殖和播散,还产生对化疗的抗性。定位参与关键代谢途径的特定酶,可能利于肝癌治疗,位点包括抑制SCD、LXRα(或称NRIH3)、ACC、IDH1或IDH2基因等。
男性的肝癌发病率高于女性。最近代谢相关研究发现睾酮会抑制白色脂肪组织中脂联素的产生,抑制AMPK信号转导,导致雄性小鼠肿瘤进展。
代谢环境还可改变肝脏免疫反应,使肿瘤细胞逃避免疫监视。此外,免疫细胞会发生代谢重编。HCC中巨噬细胞会获得M2极化,脂肪酸氧化能力增加,并利于肿瘤细胞迁移。T细胞代谢的变化影响它的定向和抗肿瘤活性;Treg细胞的扩增与其利用高能底物的能力增加。促炎免疫细胞在肿瘤微环境中会增加有氧糖酵解速率。
早期和原发性肿瘤监视
癌前状态即是特定癌基因的激活或抑癌因子的突变,诱导细胞内在(基因)或细胞外在(免疫细胞介导)监视。
基因监视。癌基因或应激驱动的恶性细胞通过细胞凋亡途径被消除,它通常由非免疫监视途径介导。最经典的途径为p53。抑制p53的基因监视功能会加速HCC发病。
免疫监视。如果原发性肿瘤的天然遗传监视失败,免疫细胞介导的监视可能会发挥作用。适应性免疫反应在具有癌变潜力的细胞监视中必不可少,它以充分的T细胞反应为标志,而这取决于肿瘤相关抗原和共刺激信号的提呈。
一般CD8+ T细胞向HCC浸润增加,预示临床结果改善。而Treg细胞在肝脏中积聚的HBV感染者,发生HCC的风险很高,这可能是由于Treg细胞会调节肿瘤特异性T细胞的功能并抑制其抗肿瘤免疫活性。在HCC患者的肿瘤微环境中发现了高比例的浸润性肿瘤CD4+CD25+FOXP3+ T细胞,这会减少CD8+ T细胞增殖并降低其功能,预示着不良预后。与此一致,Treg细胞与细胞毒性T细胞在的肿瘤内的比例有可能成为预测HCC复发和患者存活率的独立指标。但早期复发HCC中,存在高KLRB1表达的类天然CD8+ T细胞群,其抗肿瘤细胞毒性弱、克隆扩增能力低,且与预后不良相关。
NK细胞和自然杀伤T细胞是肝脏的另一道防线。与健康对照组相比,HCC患者外周血和肿瘤内组织中的肿瘤浸润NK细胞数量减少,并且NK细胞的细胞因子产生和细胞毒性功能受损。在纤维化条件下,HSCs可以通过ECM重塑以降低NK细胞活性。
同样,巨噬细胞的极化也高度异质。炎症相关巨噬细胞和浸润性单核细胞可诱导抗肿瘤反应。但在肿瘤微环境中,它们具有免疫抑制表型并通过组织重塑(如血管生成和伤口愈合)、侵袭和转移促进肿瘤进展。
肝脏微环境驱动的肿瘤转移
肝脏免疫微环境随着代谢谱的进行性变化而变化,继而可促进肝恶性肿瘤的发生。正常的肝脏内环境也可因来自其他器官恶性肿瘤细胞而发生迅速和短暂地改变使得肿瘤细胞聚集。而肝脏作为继淋巴结后最常发生转移的器官(如肠癌肝转移),主要由于存在肠-肝循环以及肝脏本身发生的免疫抑制作用。肝脏微环境的某些特定组分与肿瘤发生血液转移之间的相互作用包括以下几个方面。
肝窦内皮细胞和肝细胞共同促进肿瘤转移
恶性肿瘤释放CTC进入血液或淋巴系统,当其从血液渗出至肝脏,LSEC会出现第一道屏障,LSEC-CTC相互作用决定了后者是否形成转移性肿瘤细胞。此外,肝细胞-CRC细胞的直接相互作用,表明肝细胞可以帮助肿瘤细胞转移至肝实质。最后,代谢微环境已被证明可影响肝实质中的扩散转移。
肝脏免疫细胞形成肿瘤转移前状态
在肿瘤转移的早期阶段,KCs发挥了抗肿瘤功能。当它们的吞噬能力超负荷时,它们会改变极化并通过分泌TNF来支持CTC外渗,以增加LSECs衍生受体的表达。而HSCs的另一个转移前功能是调节免疫细胞浸润,因为它们可以吸引免疫抑制Treg细胞和MDSCs,抑制T细胞增殖和活化,从而保护肝脏中的转移细胞免受免疫细胞清除。
转移瘤的免疫监视
尽管肝脏微环境参与转移瘤的发生发展,但同时也有对转移瘤的监视机制。LSECs杀伤CTCs的同时,还对转移瘤的新生血管形成和抑制具有双重作用。宿主体内CD4+和CD8+细胞毒性T细胞对转移灶的监视在很大程度上取决于肝脏微环境。对肝脏微环境的改变,可以影响肿瘤对免疫治疗的疗效。
定位肝脏微环境
维持肝功能正常运作的复杂细胞和介质网络,营造了能维持肝脏疾病进展直至发展为肝癌的微环境。阻断该网络中的成分可能成为肝癌治疗的有效靶点。
免疫细胞的极化是肝脏微环境的关键组成部分。淋巴细胞浸润似与较低的复发风险和较好的预后相关。常规标准细胞疗法可能加速肿瘤相关抗原的释放,形成免疫防御。检测异常细胞上特有表达的抗原或识别触发前馈性肿瘤炎症循环的肿瘤相关细胞死亡途径,可能成为克服抗炎微环境和重新激活肿瘤免疫监视的新治疗武器。
将免疫疗法与多激酶抑制剂联合使用,不仅可作为二线或三线治疗,还可以用于一线抗癌治疗。它甚至有治愈肿瘤的潜力。然而不幸的是,个体对免疫治疗的反应性差异仍然是一个巨大的挑战。此外,全身治疗可能会产生不利的副作用,进一步威胁转移患者的健康。
定位能表征肝癌中异质性突变特性的特异性代谢变化,可能是增加个体对免疫疗法反应性的有效选择。此外,定位于塑造肝癌免疫微环境的代谢途径的药物可以被引入不同的联合疗法中。
结论
尽管对肝癌起源和详细分子特征的了解取得了长足进步,但能够为患者带来实质性生存和改善生活质量的治疗选择仍然很少。最近发现,组合免疫疗法可能有效促进病毒诱导的肝癌中的肿瘤监视。而非病毒病因肝癌似乎限制了免疫治疗触发的免疫监视的功效。因此,依次定位和重塑肝脏的炎症和/或代谢微环境,以及更好理解和精确靶向血管生成、纤维化和营养供应,可以增强淋巴细胞向肿瘤的浸润,从而重新建立有效的免疫监视和对免疫检查点阻断的应答。
编译:蒋龙,翁梅琳
审校:张军,缪长虹
参考文献:Li, X., Ramadori, P., Pfister, D., Seehawer, M., Zender, L., & Heikenwalder, M. (2021). The immunological and metabolic landscape in primary and metastatic liver cancer. Nature reviews. Cancer, 21(9), 541–557. https://doi.org/10.1038/s41568-021-00383-9
声明:古麻今醉公众号为舒医汇旗下,古麻今醉公众号所发表内容之知识产权为舒医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




