
本文的结果为未来研究优先效应在促进宿主-微生物积极相互作用方面可能发挥的作用提供了基础。
编译:微科盟Moon,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
大多数多细胞真核生物都拥有复杂的微生物群落,但人们对控制微生物群落组装的因素知之甚少。特定微生物的定植可能对群落组成产生长期影响,这种现象被称为优先效应。然而,单个细菌菌株对宿主微生物组的优先效应很少被研究,因此它们对微生物组功能的影响仍然未知。本研究通过实验测试了两种细菌菌株(Pseudoalteromonas tunicata D2和Pseudovibrio sp. D323)对绿藻澳洲石莼(Ulva australis)的微生物群落组装和发展的影响。
利用16S rRNA基因测序和qPCR,我们发现这两种菌株都具有优先效应,菌株D2在初始阶段引起强烈但短暂的分类变化,而菌株D323引起较弱但持续的变化。这种持续变化主要是促进性的,包括可能有益于藻类宿主的分类群。宏基因组分析表明,这些菌株对预测的微生物组功能既产生了一些相同影响(如III型分泌系统基因表达下调),也产生了某些特有影响(如抗生素抗性基因的上调)。综上,这些结果表明定植细菌对宿主微生物群落的结构和功能具有强烈的特异性影响。了解优先效应的特性是开发新型益生菌以改善宿主状况的关键。
论文ID
原名:Differential priority effects impact taxonomy and functionality of host-associated microbiomes
译名:不同优先效应影响宿主微生物群的分类和功能
期刊:Molecular Ecology
IF:6.622
发表时间:2022.1
通讯作者:Suhelen Egan
通讯作者单位:澳大利亚新南威尔士大学
DOI号:10.1111/mec.16336
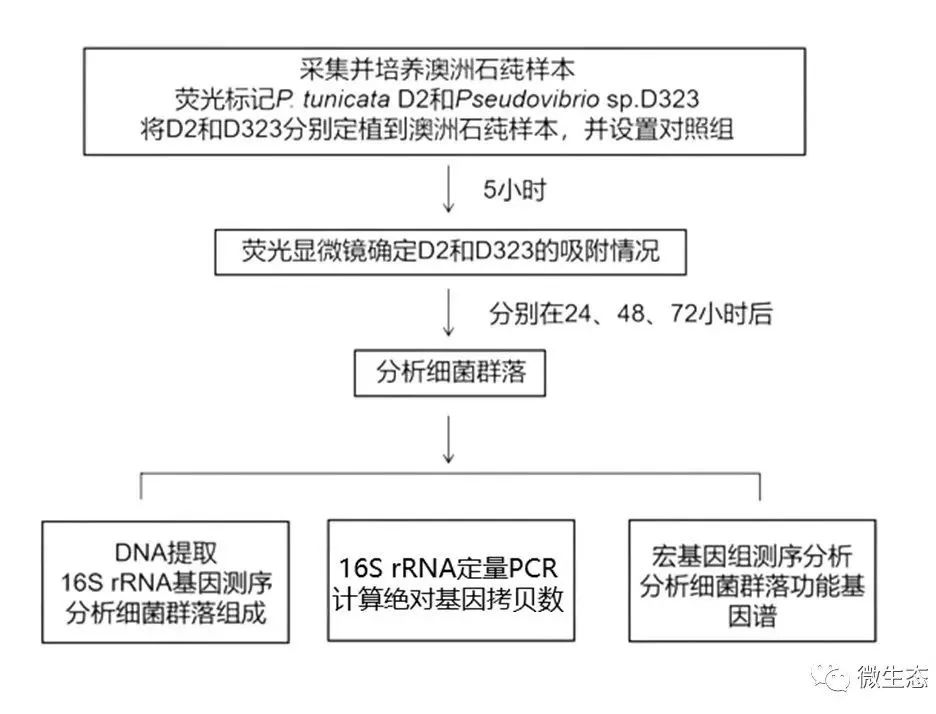
实验设计
图1 定植实验设计图。
结果
1 细菌菌株P. tunicata D2和Pseudovibrio sp. D323的定植影响藻类微生物群落的组装 首先,我们评估了D2和D323菌株成为澳洲石莼表面微生物群落成员的附着能力。我们发现,通过超声处理(见附件S1第2.1节,图S1)破坏澳洲石莼生物膜,大大提高了这两种菌株在藻类表面定植的能力(t10= -3.69;p = 0.003,图S2),这表明澳洲石莼形成的天然生物膜对入侵的细菌具有一定的抵抗力。 然后,我们评估了定植菌株是否在表面微生物群落中长期存在,并对后续群落组成和结构产生影响。为了解决这个问题,我们分析了与无定植对照组(用无菌海水培养的藻类)相比,任一菌株定植后的海藻表面微生物群落组成。对由此产生的54个样品的16S rRNA基因进行扩增子测序,产生了8,247,356条高质量reads,这些reads分属于4227个独特的序列(zOTUs)。稀疏曲线(图S3)和Good覆盖指数(99.68±0.21%;平均值±SD)表明,测序几乎完全覆盖了细菌群落。我们发现,其中三个zOTU(zOTU22、53和135)对应于菌株D2三个不同的16S rRNA等位基因,而一个zOTU(zOTU1)被鉴定为属于菌株D323,这些不纳入随后的群落水平分析。在24小时内,菌株D2定植组的序列丰度明显高于菌株D323定植组(t10 = 33.8,p = 0.05)。然而,两种菌株对应的序列丰度在48 h时下降,到72 h时,相应的zOTU之间的reads丰度没有差异(图S4)。此外,菌株D2和D323 zOTU在对照组中丰度非常低(占所有reads的0.001%至0.01%)。
用D2或D323菌株定植澳洲石莼,导致藻类微生物群落的组成和结构随着时间的推移发生显著变化(PERMANOVA,处理×时间交互作用:组成F1,2 = 2.63, p = 0.015,结构F1,2 = 1.77, p=0.027;数据S2,图2)。这种效应对群落结构的影响比对组成的影响更明显(图3)。
图2 接种D2菌株或D323菌株后,澳洲石莼的细菌群落组成(a)和结构(b)的排序。基于Sorensen(a)和Bray-Curtis(b)相似性的非度量多维尺度分析(nMDS)展示了暴露在自然海水中24、48和72小时后的微生物群落。不同的阴影表示不同组别,不同的形状表示不同时间点。
2 优先效应的程度依赖于细菌菌株
为了评估优先效应的相对影响,我们比较了菌株D2或菌株D323处理与对照组的细菌群落随时间变化的平均差异性。我们发现,与对照组相比,菌株D2处理72小时后的群落差异显著降低(D2 48h vs. D2 72h:结构F1,2 = 4.5,p = 0.03;组成F1,2 = 12.9,p = 0.005)。这与菌株D323的处理形成对比,后者在每个时间点上,群落与对照组没有任何显著性差异(p>0.05,图3,数据S2)。菌株之间的成对比较表明,在48小时内,菌株D2的优先效应程度明显大于菌株D323(即群落组成和结构与对照相比差异更大,图3,数据S2)。但是,72小时后,这种差异变得不显著。
具体而言,在用D2和D323菌株处理时,分别有276个和245个zOTU的丰度与对照组有差异(数据S3和S4,图S5和S6)。在差异丰富的zOTU中(倍数变化>2),相对于对照组,大多数在菌株D2(265)或菌株D323(245)的处理组中升高(数据S3和S4)。
图3 箱形图展示了在接种D323菌株或D2菌株24、48和72小时后,澳洲石莼的细菌群落组成(a)和结构(b)与对照组的平均差异性。D2菌株和D323菌株在每个时间点与对照组的差异值用星号表示(PERMANOVA,配对比较检验,P < 0.05)(每个时间点的n = 6)。误差条代表平均值±标准差。
3 持续和变化的优先效应
为了更好地理解所观察到的优先效应的动态特性,我们根据在单个拮抗细菌处理中与对照组相比明显上调或下调的zOTU丰度的倍数变化,进行了时间分析。这些受影响的zOTUs的丰度模式将揭示由D2或D323菌株定植引起的细菌群落组成的变化是否可能持续(长期的群落)或只是暂时的变化(可变的群落)。 许多zOTU的丰度随着时间的推移而变化,但在菌株D2或D323的处理中,分别有56和119个zOTUs保持持续的丰度,其中29个zOTUs在两个处理中变化一致(表1,数据S5)。这些常见的zOTUs大部分属于交替单胞菌(Alteromonas)、Glaciecola、Alteromonas_A和Shimia属(图4)。而每个处理组中特有的zOTUs,在菌株D323处理中发现有80%是长期存在的,而在菌株D2处理中只有19%。菌株D2处理组中的特有zOTUs来自9个属,包括Hellea、Phaeobacter、Mesoflavibacter和Pseudaminobacter等(图5a,数据S6),而菌株D323处理组的特有zOTUs分属于23个属,包括Thalassotalea、Teredinibacter、Aestuariibacter_A和Pseudoalteromonas等(图5b,数据S7)。
这些结果表明,尽管在考虑整个群落(见上文)时,菌株D323处理的优先效应相对较弱(见上文),但与菌株D2处理相比,其导致的持续变化的数量和比例相对较高。
表1 D2和D323菌株处理后,实验组和对照组之间差异丰富的序列中,随时间持续不变或变化的细菌。
图4 澳洲石莼表面建立的响应于D2和D323菌株定植的共同和持续存在的zOTUs。 热图展示了处理组和对照组之间每平方厘米16S rRNA基因绝对拷贝数有统计学差异的zOTU序列(P < 0.05)。 数字表示每个zOTU每平方厘米的绝对16S rRNA基因拷贝数。
图5 澳洲石莼表面建立的响应于D2菌株(a)或D323菌株(b)定植的特有和持续存在的zOTUs。热图展示了处理组和对照组之间每平方厘米16S rRNA基因绝对拷贝数有统计学差异的zOTU序列(P < 0.05)。所有差异丰度的zOTU都显示在(a)中;在(b)中只展示了90个zOTU中丰度最高的30个,差异丰富zOTU的完整列表可在数据S6和S7中找到。数字表示每个zOTU每平方厘米的绝对16S rRNA基因拷贝数。
4 优先效应导致细菌群落功能的菌株依赖性变化
接下来,我们分析了观察到的优先效应是否会导致功能基因的变化。虽然计算处理组和对照组之间功能谱的平均Bray-Curtis差异性时,D2处理组的群落普遍高于D323处理组,但它们的差异在统计学上并不显著(COG、KEGGS和CAZy;pairwise.adonis:P>0.05,图S7和S8)。但我们确实发现菌株D2处理组和对照组之间的基因功能差异大于菌株D323处理组和对照组之间的差异(Tukey检验,p<0.05)(例如,在总共3988个COG中,有365个受菌株D2影响,只有148个受菌株D323处理影响,表S1)。这种模式在KEGG和CAZys功能分析中也可以看到,在5318个KEGG和213个CAZys功能中,分别有354个KEGG和13个CAZys在菌株D2处理组中差异丰富,而只有136个KEGG和3个CAZys受到菌株D323处理的影响(表S1和S2,数据S8-S13)。
属于III型分泌通路的基因(COGs EscT、EscR、EscV和EscJ;以及KEGG yscU、yscT、yscR、yscQ、yscN和yscJ)在两种菌株的处理中都比对照组少(图6a、b)。此外,我们发现与对照组相比,两个菌株处理组中应激反应(hsIJ-KEGG03668,oxa-KEGG17838)、转录调控(sagS-KEGG20973,aefR-KEGG19736)、有毒化合物降解和抗药性(inhA-KEGG18199,sugE-KEGG11741)、维生素合成(ribD-COG0117)和移动遗传元件(dpo-KEGG02334)相关的基因功能均增强(图6a,b)。此外,一个属于糖基转移酶(GTs)家族的糖原或淀粉磷酸化酶编码基因(GT32)也在两个处理组中上调。
然而,大多数功能变化是单个菌株所特有的(图7,数据S12和S13),菌株D2处理组的变化数量平均比菌株D323处理高5.5倍(表2)。菌株D2处理组中上调的基因功能包括参与连接菌毛形成(trbC、traF、traG-K12059、K12057、K12056)、渗透性休克(betB-K00130)、抗生素和重金属抗性(streptogramin lyase-COG4257、scsB-K08344)、金属铁转运(cbtA),以及一个与脯氨酸亚氨基肽酶同源的基因(pip-K01295)和一个编码属于CAZys糖苷水解酶(GH)家族的α-半乳糖苷酶(GH36)的基因。菌株D2处理组中下调的基因功能包括参与sn-甘油3-磷酸转运(K05815)、铁稳态(COG4848,COG1629)、维生素K合成(MqnX)、一种假定的脂肪酶(COG1647)和两种CAZys GH158(内-β-1,3-葡聚糖酶)和GH113(β-甘露糖酶)。此外,与对照组相比,参与噬菌体(CRISPR,COG4823)、抗生素(vanX COG2173,COG4454)和次级代谢物生产(柚皮素-查尔酮COG3424)的基因在菌株D2处理组中也发生了下调(表S2,数据S8-S11)。
仅在菌株D323处理组中上调的基因包括移动遗传元件(例如:COG5527编码质粒复制蛋白的COG5527,编码噬菌体尾部和转座酶的COG3941)以及可能参与β-内酰胺酶活性(COG2367)和维生素合成(YbiA COG32236)的基因。而底物运输(腐胺、羟甲基嘧啶-K15599)和海藻酸生物合成(AlgD,K18335)相关基因则发生了下调。
图6 基于单变量GLMs分析(P < 0.05),D2 vs对照组SSW(圆圈)和D323 vs对照组SSW(三角形)之间差异丰富的COGs(a)和KEGGs(b)基因的分布。灰色和黑色符号分别代表与对照组相比下调(<0.5)和上调(>2)的基因。虚线代表倍数变化的分界线(0.5和2)。
图7 基于单变量GLMs分析(P < 0.05),D2 vs对照组SSW(a, c)和D323 vs对照组(b, d)之间前30个差异丰富的COGs(a)和KEGGs(b)基因的分布。灰色和黑色符号分别代表与对照组相比下调(<0.5)和上调(>2的基因。虚线代表倍数变化的分界线(0.5和2)。
表2 处理组和对照组之间差异丰富的COGs和KEGGs数量。
讨论
1 海藻微生物群中的菌株特异性优先效应
了解影响驱动生态群落形成的因素仍是生物领域的一个主要目标。用来解释群落形成的其中一个过程是优先效应,即一个特定物种的特征不仅决定它能否成功在该群落定植,而且还严重影响周围的群落。使用海藻澳洲石莼作为模式宿主,我们发现,用自然存在的反硝化细菌(P. tunicata D2或Pseudovibrio sp. D323)进行定植,会导致随后的细菌群落发展与对照组(即没有接种这两种菌的群落)明显不同。这些观察到的优先效应的性质与菌株有关:菌落发展前期,菌株D2存在下的群落变化比菌株D323更明显。 虽然随着时间的推移和菌株D2丰度的降低,群落开始类似于对照组细菌群落,但群落结构仍然与对照组明显不同,甚至当菌株D2丰度接近于零的时候也是如此(图S4)。
这表明,菌株D2的优先效应具有遗留效应,并通过一些初始(相互)作用/影响受体微生物群落的形成过程。相比之下,菌株D323的丰度随着时间的推移保持一致,这导致对群落的优先效应相对较小,但持续时间更长。 菌株D2和D323之间优先效应的差异可以用两个物种之间的性状差异来解释,例如它们产生的次级代谢物的数量和类型的差异,它们利用特定碳水化合物来源和/或获得微量元素的能力。这些性状对直接或间接改变环境条件的相对贡献可以进一步解释优先效应的差异。直接使用物种特征来预测优先效应的强度还没有得到广泛的研究,在这个领域的进一步研究可以提高我们对自然界和工程系统中微生物群落动态的理解。
2 持续存在的优先效应主要是有利的
本研究观察到,与菌株D2相比,菌株D323对附生群落产生一致zOTU变化的比例更高(分别为49%和20%,表1)。其中一部分(即菌株D2约为51%和菌株D323约为24%,表1)是两个菌株所共有的,包括属于Alteromonas、Glaciecola和Shimia的zOTU增加。这些属的成员通常与大型藻类共生,并被认为是溶解性有机物的一般消费者,能够快速响应有机碳的流动。
例如,以前的研究已经确定Glaciecola spp.是硅藻水华早期阶段海藻衍生有机碳的主要消费者。未来的研究将需要确定,在暴露于D2或D323的群落中,这些细菌属的较高丰度是否与这些菌株从石莼生物质中释放DOM的能力有关(直接或间接)。 相对于对照组,细菌定植处理组表现出相对较少的zOTU丰度下降(11个zOTU,数据S3和S4),并且全部来自D2定植组。减少的zOTU主要属于Alphaproteobacteria和Bacteroidia,包括那些属于Lewinella和Maribacter属的成员,它们能够产生类胡萝卜素和分解复杂聚合物。
虽然菌株D2的竞争性排斥可能是这一结果的基础,但值得注意的是,菌株D2确实产生了一些具有生物活性的次级代谢物,可能有助于直接抑制作用。事实上,D2(还有D323)的抗菌特性预计会对更多的群落成员产生负面影响,但这在此项研究中并没有被发现。菌株D2处理组的大多数zOTU导致群落分类群的增加,其中一些以产生抗氧化剂(Flavobacteriaceae)或自身产生抗菌剂(Phaeobacter)而闻名,表明菌株D2可能促进有益细菌的建立。 本研究观察到,在D323菌株存在下形成的群落中独特富集的zOTUs数量高于菌株D2。这些结果表明,虽然菌株D2的存在导致群落在初期与对照组相比差异更大,但菌株D323的定植会使群落发生持续时间更长的变化。
具体来说,菌株D323的独特富集变化包括属于Thalassotalea、Pseudoalteromonas、Teredinibacter和Aestuariibacter_A的zOTUs。研究人员研究了其中每一种作为复杂多糖降解者和/或次级代谢物生产者的作用。Teredinibacter的成员是船蛆的内生菌,在纤维素降解中发挥重要作用,但也被发现可以固氮并为其无脊椎动物宿主提供维生素。Aestuariibacter是最近报道的一个与Alteromonas相近的属,虽然对这类菌了解不多,但它们在商业栽培的Caulpera的健康株中含量丰富,有人猜测它们也可能对其他绿色海藻发挥有益作用。有趣的是,观察到的优先效应主要导致系统发育上与定植菌株无关的菌株的增加。群落生态学预测,当存在最小的生态位空间竞争时就会出现物种共存,宏观生态学领域预测这一特征与系统发育距离密切相关。虽然这一理论应用于微生物群落形成的程度目前还存在争议,但这里观察到的优先效应可能部分是由于新来的或已经居住的细菌占据了与D2和D323菌株不同的生态位的结果。
3 优先效应表现在产生的附生群落的功能差异上
微生物生态学中一个备受争议的假设是,系统发育距离反映了功能的差异。然而,看似系统发育不同的微生物群落可以有高度的功能相似性。这种功能上的相似性表明,群落形成主要是由其中成员的功能而不是他们是谁(分类学)驱动的。在这里,由于任一菌株的定植而导致的分类学变化对群落的预测功能有影响。例如,D2或D323菌株的定植导致编码III型分泌系统(T3SS)多种成分的基因下调,这些基因在致病菌中广泛存在,其功能是将各种毒素和其他毒力效应物直接运送到宿主细胞中。虽然还需要进一步研究,但这一观察结果表明,在有D2或D323菌株存在的情况下,建立的微生物群的致病潜力降低。其余的常见功能在任一菌株定植后都得到了上调,包括参与环境压力(如重金属/药物外排泵和热休克蛋白)和移动遗传元件(如转座酶和噬菌体蛋白)的功能,表明一个更具弹性的微生物群具有增加遗传交换的能力。
有趣的是,虽然与菌株D323相比,菌株D2的优先效应导致所产生的群落分类学特征的持续变化较少,但在考虑这些细菌群落的预测功能时,我们观察到了相反的趋势;即相对于对照组,菌株D2的定植导致了比菌株D323更大数量的特有功能基因变化。这些模式表明,菌株D2对具有不同功能特征的群落成员有更大的影响,因此,可能解释了观察到的菌株D2的遗留效应。 在接种了D2菌株的群落中,独特富集的基因与质粒转移、对链霉素抗生素的耐药性、渗透性休克和重金属耐受有关。菌株D2的处理还导致了与次级代谢物的生产有关的功能的下调。黄酮类化合物,包括柚皮素,主要是植物代谢物,但最近在放线菌中发现其具有抗菌活性并保护细胞免受紫外线和渗透压的影响。
作为一种潮间带海藻,澳洲石莼经常受到干燥的影响,虽然这些差异丰富性状的净影响尚不清楚,但由于菌株D2的优先效应,抵抗紫外线和渗透胁迫的能力降低,可能导致微生物群和宿主对这种环境生态位的适应性降低。 在接种D323菌株的群落中,独特富集的基因包括编码β-内酰胺类抗生素抗性和核黄素(维生素B2)生物合成途径的基因。核黄素已被证明在植物的氧化应激、耐旱性和抵御病原体方面发挥作用,将其添加到小球藻的培养物中可促进生长和色素产生。也有人认为核黄素交换可能是共生系统的一个共同特征。相反,与机会致病菌有关的特征,如腐胺的运输和胞外多糖海藻酸盐的产生,在菌株D323存在下建立的微生物群中发生了下调。综上所述,这表明菌株D323的优先效应有可能导致的微生物群特征变化整体上有利于宿主。然而,必须说明的是,需要进一步的研究来确认菌株D2和D323的优先效应是否也会影响宿主的健康。
综上所述,本研究证明了特定细菌的定植可以对宿主微生物群的后续发展产生深远和不同的影响。这些观察到的优先效应似乎不是功能等效类群的简单替代,而是导致群落功能潜力的明显差异。尽管两个菌株都有拮抗特性,但优先效应的程度和类型与菌株有关。虽然菌株D2在初期对群落有更大的影响,但菌株D323在群落中产生了更持久的变化,包括对藻类宿主有潜在好处的功能。到目前为止,微生物优先效应对宿主健康的影响主要集中在负面作用上,本文的结果为未来研究优先效应在促进宿主-微生物积极相互作用方面可能发挥的作用提供了基础。这不仅将促进我们对群落生态学的基本理解,而且将为现有(如人类健康和农业)和新兴应用开发新的益生菌带来更广泛的好处,包括增强微生物组以提高宿主对环境压力的适应性。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您









































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




