
本研究结果强调了细菌生活史策略和成员关系在自然环境中决定其繁殖成功方面的重要作用,揭示了环境选择和生物因素在塑造细菌群落中的基本耦合。
点击蓝字↑↑↑“微生态”,轻松关注不迷路
生科云网址:https://www.bioincloud.tech
编译:微科盟沐木,编辑:微科盟居居、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读 自然微生物群落普遍存在营养缺乏问题,影响物种繁殖和共存。但目前尚不清楚是否存在生物和非生物因素影响微生物物种丰度的一般规律。本研究表明,核糖体RNA基因操纵子(rrn)拷贝数是一种与细菌生长速率和营养需求相关的基因组特征,在营养丰富的沿海沉积物中从丰富生物圈到稀有生物圈表现为减少,但在营养稀缺的全球海洋的远洋带中却表现出相反的模式。这两种模式都以群落水平rrn拷贝数和养分之间的正相关为基础。此外,与海洋水样相比,在沿海沉积物中观察到更多由负网络关联推断的物种间共排斥。养分操纵实验产生了养分有效性对rrn拷贝数和网络关联的影响,这与我们的实地观察结果一致。基于这些结果,我们提出了一个“饥饿游戏”假设,使用rrn拷贝数、生态相互作用和养分可用性来定义微生物物种丰度规则。
论文ID
原名:Nutrient supply controls the linkage between species abundance and ecological interactions in marine bacterial communities
译名:养分供应控制海洋细菌群落中物种丰度与生态相互作用之间的联系
期刊:Nature Communications
IF:17.694
发表时间:2022.1
通讯作者:温东辉,杨云锋
通讯作者单位:北京大学环境科学与工程学院;清华大学环境学院
DOI号:10.1038/s41467-021-27857-6
实验设计
结果
1 丰富、中间和稀有生物圈中单个OTU的rrn拷贝数 根据OTU的丰度和出现频率对沿海沉积物和海水样品中的丰富、中间和稀有生物圈进行了分类(详见图1)。数据集中不到1.46%的OTU被归类为丰富生物圈,而61.40%~98.51%的OTU归为稀有生物圈,其余的OTU处于稀有和丰富生物圈之间的中间生物圈(补充表1)。 根据rrnDB数据库估算了细菌OTU的rrn拷贝数。大多数沿海沉积物和海水群落的OTUs rrn拷贝数在丰富、中间的和稀有生物圈之间存在显著差异(图1b和补充表1)。除地中海沿岸和悉尼沿岸沉积物外,OTU的rrn拷贝数在大多数沿海沉积物中从丰富生物圈到稀有生物圈表现为减少,但在全球海水中呈上升趋势(图1b和补充表1)。与沿海沉积物相比,海水中丰富生物圈的OTUs rrn拷贝数显著降低(补充图1a),但稀有生物圈的OTUs rrn拷贝数显著增高(补充图1b)。沿海沉积物中丰富生物圈一般包括芽胞杆菌目(6~11个rrn拷贝;杭州湾和使命湾)、梭菌目(5个rrn拷贝;地中海沿岸)、脱硫杆菌目(4个rrn拷贝;普利茅斯港、悉尼沿海和地中海沿岸)、交替单胞菌目(5个rrn拷贝;墨西哥湾和悉尼沿海)、红杆菌目(4个rrn拷贝;地中海沿岸)、黄杆菌目(3个rrn拷贝;悉尼沿海)、红螺菌目(4个rrn拷贝;墨西哥湾)和海洋螺菌目(5个rrn拷贝;墨西哥湾)。在全球海水中,属于丰富生物圈的105个OTU中有71个是只有一个rrn拷贝的SAR11。为了评估SAR11和其他具有单个rrn拷贝的OTU对rrn拷贝数的影响,我们从数据集中删除了这些OTU。我们发现海水中丰富生物圈的rrn拷贝数仅显著低于使命湾和墨西哥湾的沉积物(补充图1c)。相比之下,稀有生物圈的数量仍显著高于沉积物样品(补充图1d),表明海水中的单个rrn拷贝OTU对丰富生物圈的低rrn拷贝数贡献最大。
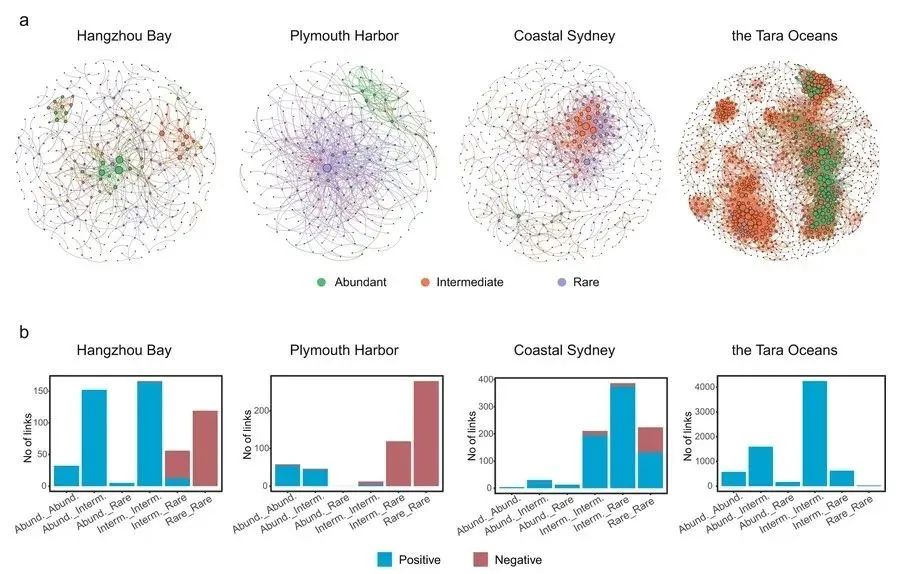
图1 沿海沉积物和全球海水中细菌OTU的rrn拷贝数。a 在本研究中调查的沿海沉积物和海水细菌群落的位置图。b 丰富、中间和稀有生物圈中OTUs rrn拷贝数。OTUs根据其相对丰度和出现频率分类:(i)丰富生物圈,在>50%样本中OTUs相对丰度≥0.1%,并且在80%以上样本中出现;(ii)稀有生物圈,所有样本中OTU的相对丰度<0.1%;(iii)中间生物圈,属于稀有生物圈或丰富生物圈以外的OTU。条形表示平均值,误差条表示标准误差。样本量和详细的汇总统计数据列在补充表1中。当P<0.001时,单因素方差分析的未校正P值随LSD测试标记为***,当P<0.01时标记为*,当P<0.05时以及当P>0.05(不显著)时标记为ns。在Dois Rios Estuary的数据集中,没有OTU被归类为丰富生物圈,因此该栏被隐藏。c,d 沿海沉积物和海水及海水细菌群落的群落水平rrn拷贝数和系统发育多样性。条形上方的小写字母表示显著差异(P<0.05,单因素方差分析LSD检验)。图(c和d)中的样本量如下:使命湾,20;杭州湾,72;墨西哥湾,6;普利茅斯港,66;地中海沿岸,11;悉尼沿海地区,60;Dois Rios Estuary,9;塔拉海洋,139。 2 群落水平的rrn拷贝数 如先前的研究所述,我们将群落水平rrn拷贝数计算为每个群落成员的丰度加权平均rrn拷贝数。全球沿海沉积物群落水平的rrn拷贝数平均为2.74±0.06,范围为1.59-7.61。使命湾(3.52±0.25)、杭州湾(3.18±0.17)和墨西哥湾(2.79±0.06)高于其他沉积物。全球海水中群落水平的rrn拷贝数在深海中高于表层海水,这可能是由于深海中SAR11丰度较低(补充图2)。然而,它的平均拷贝数为1.54±0.01,范围在1.39-2.53之间(图1c),仍显著低于沉积物(P<0.001,ANOVA)。未加权群落水平rrn拷贝数在海水中的丰度也低于在沉积物中的丰度(补充表1),这表明忽略分类单元丰度不会影响我们的结果。通过计算OTUs在系统发育树上的平均成对距离(MPD),我们发现群落多样性最高的群落(如图1d中地中海沿岸和悉尼沿海)并没有最高或最低的群落水平rrn拷贝数(图1c),表明这些群落的OTUs覆盖了更广泛的系统发育谱系。 群落水平的rrn拷贝数与沿海沉积物和海水样品的所有测量营养物质(r=0.410~0.782,P<0.013,皮尔逊相关性)呈正相关(补充表2)。当控制潜在系统发育结构的差异时,Partial Mantel检验证实了群落水平rrn拷贝数和营养物质之间的显著联系(r=0.130~0.602,P<0.029)(补充表3)。在计算基于丰富、中间和稀有生物圈的rrn拷贝数时,我们发现它们与营养物质的相关性变化很大(补充表2)。在杭州湾,丰富和中间生物圈的rrn拷贝数与N和P的正相关性强于稀有生物圈。相比之下,全球海水中稀有和中间生物圈的rrn拷贝数与N和P的相关性比丰富生物圈更显著(补充表2)。 3 网络分析 为了评估群落中细菌成员之间潜在的生态相互作用,我们为杭州湾、普利茅斯港和悉尼沿海沉积物以及全球海水中的细菌群落生成了基于随机矩阵理论(RMT)的关联网络(图2a),因为这些数据集有足够的样本量(至少60个)允许进行可靠的网络分析。所有网络都表现出复杂系统的典型特性,如无标度、小世界和模块化特征。它们显示出与其相应随机网络显著不同的拓扑指数,表现出非随机特征(补充表4)。 计算了每个网络的正负关联(即网络连接),以检查物种间的共存和共排斥模式。我们观察到沿海沉积物中细菌群落的负相关比例(杭州湾,31.13%;普利茅斯,80.35%;悉尼沿海,14.38%)比全球海水(0.04%)高得多(图2b)。尽管沿海样品的细菌群落组成不同,但沿海沉积物中的大多数负相关分布在稀有生物圈内或稀有和中间生物圈之间(图2b)。与此形成鲜明对比的是,丰富生物圈内的关联主要是正相关。
图2. 沿海沉积物和全球海水中细菌OTU的网络关联。a 网络代表基于随机矩阵理论的相关模型,该模型源自每个数据集的所有样本,其中节点代表OTU,节点之间的链接代表显著相关性。节点大小与其程度成正比,颜色表示丰度类别(即丰富、中间和稀有生物圈)。网络分析的样本量如下:杭州湾,72;普利茅斯港,66;悉尼沿海地区,60;Dois Rios Estuary,9;塔拉海洋,139。b 根据相关OTUs的丰度类别对网络链接进行总结。 4 验证养分供应对细菌群落的影响 我们通过微观实验控制杭州湾收集的沉积物中的NH4+-N和PO43--P浓度,研究了养分有效性对细菌群落组装的影响。具体来说,我们添加了5 mg L-1 NH4+-N和0.5 mg L-1 PO43--P(表示营养供应不足),以及50 mg L-1 NH4+-N和5.0 mg L-1 PO43--P(表示高营养供应),在微观对照中没有添加营养。在开始三天,所有微观实验中的细菌多样性都大大减少,但随着时间的推移,在分类学和系统发育上又逐渐恢复(补充图3)。28天后,低和高营养供应下的细菌多样性相当(P=0.12),但显著(P<0.05)高于微观实验对照(补充图3)。基于Bray-Curtis距离的主坐标分析(PCoA)揭示了养分供应条件下细菌群落的不同演替轨迹(补充图4),时间和营养供应均具有显著效应(P<0.001,Adonis)(补充表5)。
在三种养分供应条件下使用与图1相同的标准将OTU分配到每个细菌群落的稀有、中间和丰富生物圈中(补充表6)。对照微观世界中丰富和中间生物圈的rrn拷贝数相当(P=0.61),但高于稀有生物圈(P<0.001,图3a)。在低和高营养供应条件下,rrn拷贝数由丰富、中间到稀有生物圈减少(所有比较中P<0.05,图3a)。群落水平的rrn拷贝数与营养物质之间的相关性强度随着养分供应的增加而显著增加(补充表7),这一趋势在丰富生物圈中比稀有生物圈更加显著(图3b)。我们构建了对照、低、高养分供应样本的个体关联网络。与对照组相比,补给养分时OTUs的联系更为紧密(图4),平均联结度(补充表8)和负相关比例(补充表9)更高。在3个网络中,稀有生物圈OTUs的负相关比例超过一半(60.7~73.6%)。除了少数例外,大多数相关OTUs对(即丰富-丰富,丰富-中间,丰富-稀有和中间-稀有)的负相关比例随着养分供应的增加而增加(补充表9),表明养分供应导致群落成员之间的负相关。
图3 养分供应对rrn拷贝数的影响。a微观实验中丰富、中间和稀有生物圈中细菌OTU的平均rrn拷贝数。条形表示平均值,误差条表示标准误差。当P<0.001时,将单因素ANOVA的未校正P值标记为***,当P<0.01时标记为**,当P<0.05时标记为*,当P>0.05时标记为ns(不显著)。样本量(即OTU的数量)列在补充表6中。使用与图1相同的标准,根据其在28天内的丰度变化对OTU进行分类。b热图显示了细菌丰富、中间和稀有生物圈的养分浓度和群落水平rrn拷贝数之间的Pearson相关系数。Pearson相关系数的P值(n=15,每个相关测试的生物学独立样本)在P<0.001时标记为***,当P<0.01时标记为**,当P<0.05时标记为*。
图4 养分供应对细菌影响的网络关联分析。a网络数据来源于从三种养分操作实验中收集的时间序列样本(0、3、7、14、28天):无供应(对照)、5 mgL-1NH4+-N和0.5 mgL-1PO43--P供应(低),50 mgL-1NH4+-N和5.0 mgL-1PO43--P供应(高)。节点颜色表示OTUs的丰度类别,节点大小与其在网络中的度成正比。b箱线图表示属于不同丰度类别的OTUs的节点度。箱线图的样本量(即单个OTUs数量)如下:对照中,丰富n=60,中间n=75,稀有n=901;低供应中,丰富n=77,中间n=94,稀有n=1130;高供应中,丰富n=75,中间n=87,稀有n=1166。
讨论
对全球海洋细菌群落的分析允许探索在个别研究中可以观察到的广义生态规则。本研究表明,rrn拷贝数、环境养分供应和潜在生态相互作用的细菌基因组特征是海洋细菌群落组装的核心驱动因素。通过考虑rrn拷贝数的一般细菌基因组特征,并结合当地环境养分供应以及潜在的生态相互作用,我们提出了一个解释海洋细菌群落组装和分类群丰度的假设。我们通过结合几种现有的理论/假设(如环境选择理论、生长速率假设、进化博弈论和生态位构建理论)来主张这一假设,提出我们称之为“饥饿游戏”的假设,以突出潜在生态相互作用和营养供应在塑造细菌群落中的相互作用。我们的假设包含两个规则:(1)天然细菌群落的物种丰度谱受基因组性状rrn拷贝数的影响,其中高rrn拷贝数有利于富营养环境,而低rrn拷贝数有利于贫营养环境;(2)天然细菌群落以合作为主,但在富营养环境中竞争更为频繁。我们生成一个概念图来总结它(图5)并在下面详细阐述了“饥饿游戏”假设。
图5 “饥饿游戏”假设的概念图。天然细菌群落中的物种丰度由rrn拷贝数、生态相互作用和养分有效性的基因组特征共同决定。营养稀缺的环境有利于rrn拷贝数低的物种,而营养丰富的环境有利于rrn拷贝数高的物种。在营养稀缺的环境中,促进合作,例如从代谢物交换中互惠互利,以缓解资源限制。相比之下,在营养丰富的环境中,物种之间存在更多的生态相互作用,其中竞争发挥了更大的作用。 (1) 营养丰富的环境有利于细菌的快速生长,而营养缺乏则会选择出营养利用效率高的细菌。 在纯培养物中,研究表明rrn拷贝数是细菌适应养分有效性的可靠指标。本研究中,我们表明在自由生活的海洋细菌群落中也是如此(图1、3和补充表)。然而,与颗粒相关的微生物群落具有完全不同的模式,其中诸如交替单胞菌目、海洋螺旋菌目、黄杆菌目、和红螺菌目等富营养菌更为丰富,它们比自由生活的微生物具有更高的生长速率和rrn拷贝数,并且在沿海沉积物中也很常见。这类似于深海水中寡营养生物(例如SAR11)的减少和富养生物的富集(补充图2),其中由于颗粒沉积,有机物质的可用性更高。此外,稀有、中间和丰富生物圈的rrn拷贝数不同,这取决于营养条件(图1b、3a)。这一发现通过将rrn拷贝数确定为与物种繁殖成功相关的基因组特征,推进了稀有和丰富生物圈的生命史策略假设。由于核糖体富含P,因此核糖体产量高的生物体的生物量C:P和N:P比率较低。因此,rrn拷贝数可以解释增长率假设,因为不同的rrn拷贝数在生物化学计量方面有所不同,影响细菌资源需求和生长潜力。 尽管高rrn拷贝数与沿海沉积物中丰富生物圈和海水自由生活部分中的稀有生物圈有关(图1b),但群落水平的rrn拷贝数与营养物质之间存在一致的正相关关系(补充表2和S5)。沿海沉积物中的群落水平rrn拷贝数为2.74±0.06,高于土壤和植物相关微生物群落(2.2,基于地球微生物组计划),但低于动物相关样本(3.4)。所有这些数字都高于海水中的数字(1.54±0.01,图1c),并且与它们的营养水平一致,这增加了使用群落水平rrn拷贝数来预测生态建模中环境的生物营养可用性。然而,具有不同生活史策略的不同系统发育谱系可能具有相同的rrn拷贝数,因此在将群落水平的rrn拷贝数与营养可用性联系起来时,应考虑系统发育结构的影响。 (2)复杂的生态相互作用导致微生物群落高度动态的自组织,称为进化博弈。在自然细菌群落的成员中,争夺稀缺食物和有限的生存空间是很常见的。饥饿游戏策略盛行,倾向于根据食物供应增加物种合作或竞争。有限的营养供应减少了次生代谢物的分泌,这导致利用次生代谢物的群落成员的食物短缺。合作可以提高生产力,因为互惠互利的物种可以参与分工和交换基本代谢物,从而充分利用养分。相反,成员之间的竞争可以相互制衡,从而对群落产生稳定作用。 当养分充足时,富营养细菌会胜过贫营养细菌,反之亦然。出乎意料的是,丰富的分类群之间的网络关联在沉积物和海水样品中主要是正相关(图2b和补充表9),这可能是由于竞争排斥引起的优势物种的稳定共存和生态位划分的平衡。贫营养海水中的低细胞密度降低了相遇频率,阻止了生态相互作用。然而,代谢物交换对于具有小而简单的基因组的物种(如SAR11)尤其重要,因为由于基因组减少需要代谢外包。在营养缺乏的情况下,合作生长是首选(图2b和补充表9),是减轻多样性损失和增强稳定性的合理机制。当有更多的营养可用时,活跃的细菌生长可能会产生有害代谢物并改变环境,从而抑制群落中竞争微生物的生长。获胜物种的“自私”阻碍了物种的共存,从而减少了生物多样性。积极的生长也可能导致对其他必需营养元素的竞争,抵消强大竞争对手产生的生命威胁。营养物质的选择可以随着消耗而减少,最初的赢家产生的代谢物可以为其他微生物开辟新的生态位,反过来提供新的代谢物,支持更广泛种群增长,从而导致群落多样性的恢复(补充图3)。因此,养分供应和代谢物分泌同时通过不同的路径推动群落演替。然而,养分可用性对群落稳定性的影响尚不明确,因为较低的生物多样性会降低稳定性,但在自然和微观群落中观察到的养分改良导致的较高的负网络关联可能会增强稳定性(图2b和补充表9)。 然而,人们应该明白只有当生态相互作用持续且适当地影响物种丰度并能通过适当的统计方法检测到时,从关联网络推断生态相互作用才是可行的。迄今为止,由于所有的实验室实验都局限于合成或简单的群落,因此如何检测自然环境中的实际生态相互作用仍然是一个棘手的问题。因此,关联网络分析仍然是推断潜在生态相互作用的主要工具。通过关注网络属性的聚合模式和网络之间的比较,我们可以减少大型数据集中典型的高噪声引起的假阳性和阴性的影响。 综上所述,通过比较沿海沉积物中的细菌群落和海水中自由生活部分的细菌群落,我们得到的证据表明,微生物物种丰度分布可以通过养分有效性、rrn拷贝数和生态网络来解释。本研究结果强调了细菌生活史策略和成员关系在自然环境中决定其繁殖成功方面的重要作用,揭示了环境选择和生物因素在塑造细菌群落中的基本耦合。 原文链接: https://www.nature.com/articles/s41467-021-27857-6 获取此篇微文原文pdf请扫描下方二维码联系微科盟多组学老师即可。
微文推荐阅读
土壤微生态文献包免费领取 | 150篇近两年高影响因子土壤微生态相关文章
根际微生态文献包免费领取 | 60篇近两年高影响因子植物根际微生物相关文章
肠道微生态文献包免费领取 | 100篇近两年高影响因子肠道菌群与消化道疾病相关文章
元素循环微生态文献包免费领取 | 60篇近两年高影响因子植物-土壤碳氮磷相关文章
获取此文献原文PDF、申请加入学术群,联系您所添加的任一微科盟组学老师即可,如未添加过微科盟组学老师,请联系组学老师45,无需重复添加。
了解更多菌群知识,请关注“微生态”。
点击阅读原文,直达原文网站,如需原文PDF可联系组学老师获取
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




