

图鉴!癌症代谢解析




1.自从2000年Hanahan和Weinberg发表了“The Hallmarks of Cancer“以来,我们对癌症了解得越来越深入。在过去的20多年里,癌症的标志已从最开始的6个扩充到今天的14个。其中在2011年添加的一个标志是重编程的细胞代谢。我们今天就来聊一聊癌症代谢。图源:7

2.细胞代谢最重要的一个过程是有氧呼吸。细胞最主要的能源燃料是葡萄糖。在有氧的条件下,细胞通过一系列的反应把葡萄糖分子降解成水和二氧化碳,并将分子中储存的能量“变现”为ATP。ATP是身体的能量货币。每个葡萄糖分子完全降解后可产生36个ATP分子。细胞代谢就是由若干个类似于有氧呼吸的代谢途径组成,提供、储存和消耗能量,降解和合成大分子,维持细胞的结构和功能,帮助细胞生长、繁殖,甚至凋亡。图源:21

3. 这是人体细胞代谢的全景图。红线圈出的部分是葡萄糖降解的完整途径。这一过程又分为两个阶段:发生在细胞质中的、葡萄糖转化为丙酮酸的糖酵解(glycolysis)过程(图中红色背景部分),和发生在线粒体中的三羧酸循环(TCA cycle)和氧化磷酸化(Oxidative phosphorylation或OXPHOS)(红圈中香槟色背景部分)。想更深入了解细节的朋友请登录metabolicpathways.stanford.edu网站下载高清图。这个图给我的总体印象是,细胞代谢太复杂了:剪不断,理还乱,只好无言独上西楼。图源:22

4.面对复杂的代谢网络,我们可以先由简入手。在内因(原癌基因和肿瘤抑制基因的突变)和外因(肿瘤微环境或TME)的作用下,肿瘤细胞修改了核心代谢程序以此来适应细胞快速增殖的三个基本需求:快速生成ATP以维持能量状态(bioenergetics);增加大分子的生物合成(biosynthesis);加强维持适当的细胞氧化还原状态(redox homeostasis)。分析、研究癌症的代谢离不开这三大版块。图源:adapted from 25

5. 人类研究和利用癌症代谢已经跨越百年,其中有几个重要的里程碑:
a. 1923-1927期间,德国生理学家Otto Warburg发现肿瘤细胞在有氧的条件下也利用糖酵解,大量摄取葡萄糖,把它们转化为丙酮酸后,丙酮酸不进入线粒体,而是再变成乳酸。这样,体内的葡萄糖往往富集在肿瘤处,而肿瘤组织中又有高浓度的乳酸。由此这种现象被称为Warburg效应。
b. 1948年,Sidney Farber 在《新英格兰医学杂志》上发表了一篇开创性论文,描述了抗叶酸药物氨基蝶呤缓解了儿童患者的急性白血病。氨基蝶呤抑制了二氢叶酸还原酶,阻断了核酸和蛋白的合成,成为一大类化疗药的鼻祖。
c. 90年代,致癌基因、肿瘤抑制因子和新陈代谢之间的联系开始被发现,引发了对癌症代谢的重新关注。
d. 近10年来,免疫疗法的成功凸显了肿瘤免疫微环境 (TIME) 对有效抗癌治疗策略的重要性。而肿瘤细胞、免疫细胞、基质细胞及其它类细胞的代谢活动和产物对形成肿瘤微环境起到了重要作用。
e.在2017-2019期间,两种突变型异柠檬酸脱氢酶(mutIDH) 抑制剂 Enasidenib (商品名Idhifa) 和 Ivosidenib (商品名Tibsovo)先后获得 FDA 批准,用于治疗 IDH 突变型复发或难治性急性髓系白血病 (AML)。

6. 咱们先看一下Warburg效应。我们一提起癌症代谢,首先想到的是Warburg效应。在大部分正常组织中,一个葡萄糖分子通常从头到尾走完降解过程——糖酵解、三羧酸循环和氧化磷酸化。这样葡萄糖分子可以最大效率地释放能量——一个分子可以产生36个ATP分子。 在无氧的条件下,葡萄糖转化为丙酮酸再变成乳酸,只产生两个ATP。而在肿瘤组织中,不管在有氧还是无氧条件下,糖酵解占据了主导地位。有氧糖酵解不是肿瘤细胞所独有的。某些快速繁殖的正常细胞也依赖于这一过程,这常常使得用靶向该通路来治疗癌症的策略在实施中易引起安全性的问题。图源:18

7. 相对于正常细胞(图a),肿瘤细胞(图b)向有氧糖酵解的转变是由多种致癌信号通路驱动的:
a. PI3K激活AKT,AKT通过直接调节糖酵解酶和激活mTOR 来刺激糖酵解。
b. 肝激酶B1 (LKB1) 肿瘤抑制因子通过激活AMPK、抑制mTOR 来抵制糖酵解表型。而在肿瘤细胞中,这条通路往往被切断。
c. mTOR以多种方式改变新陈代谢,其中一种方式是通过增强缺氧诱导因子1 (HIF1) 活性。
d. HIF1启动缺氧适应性转录程序,包括增加葡萄糖转运蛋白(GLUT)、糖酵解酶和丙酮酸脱氢酶激酶 (PDK) 的表达,从而阻止丙酮酸进入TCA循环。
e. MYC与HIF 合作激活几个编码糖酵解蛋白的基因,但也会增加线粒体代谢。
f. 肿瘤抑制因子 p53 通过诱导TIGAR来抑制糖酵解,从而对抗糖酵解表型,此外,p53还通过SCO2增加线粒体代谢和支持 PTEN 的表达。p53往往在肿瘤中失活,其突变频率高于LKB1。
g. OCT1(也称为POU2F1)激活相应基因的转录,驱动糖酵解,并抑制氧化磷酸化。
糖酵解是一个非常浪费的过程——与走完线粒体氧化磷酸化的全过程相比,葡萄糖变成乳酸只能获得1/18的ATP,但产生ATP的速度却提高了近100倍。癌细胞用浪费换来了速度,迅速供能和合成。就好像两个人比赛吃切好的西瓜,正常细胞老老实实地把一块西瓜的瓤都啃光了再去拿下一块,而癌细胞每块西瓜只咬顶尖一口,随手一扔又去抓下一块。癌细胞不守规则疯狂地掠夺西瓜,让别的细胞无瓜可吃。图源:25

8.近些年来我们透过Warburg效应看到了更深层次的复杂性。以前认为糖酵解/OXPHOS是一个开关,癌细胞完全拧到了发酵档而关闭了OXPHOS(图A)。现在发现,癌细胞中的OXPHOS同样活跃(图B)。肿瘤细胞只是几乎关闭了糖酵解与OXPHOS之间的渠道。TCA循环提供了大分子(核酸、脂类、蛋白)合成的起始原料,是癌细胞迅速繁殖不可或缺的。但大部分来自葡萄糖的丙酮酸分子被引入歧途,无法进入线粒体驱使TCA循环转动。癌细胞又靠什么来驱动TCA循环呢?癌症代谢中的第二主角谷氨酰胺闪亮登场。图源:5

9. 谷氨酰胺是一种非必需氨基酸,可在体内合成。一个往往被忽视的现象是,在体外细胞系的培养液的配方中,谷氨酰胺的浓度是其它氨基酸的十倍甚至百倍。体外细胞系往往是肿瘤细胞或类肿瘤细胞衍生来的,而适合它们的培养液的配方是经过多次实验摸索出来的。肿瘤细胞为什么会渴望谷氨酰胺呢?这是由它在代谢途径网络的枢纽地位决定的。图源:14, 23
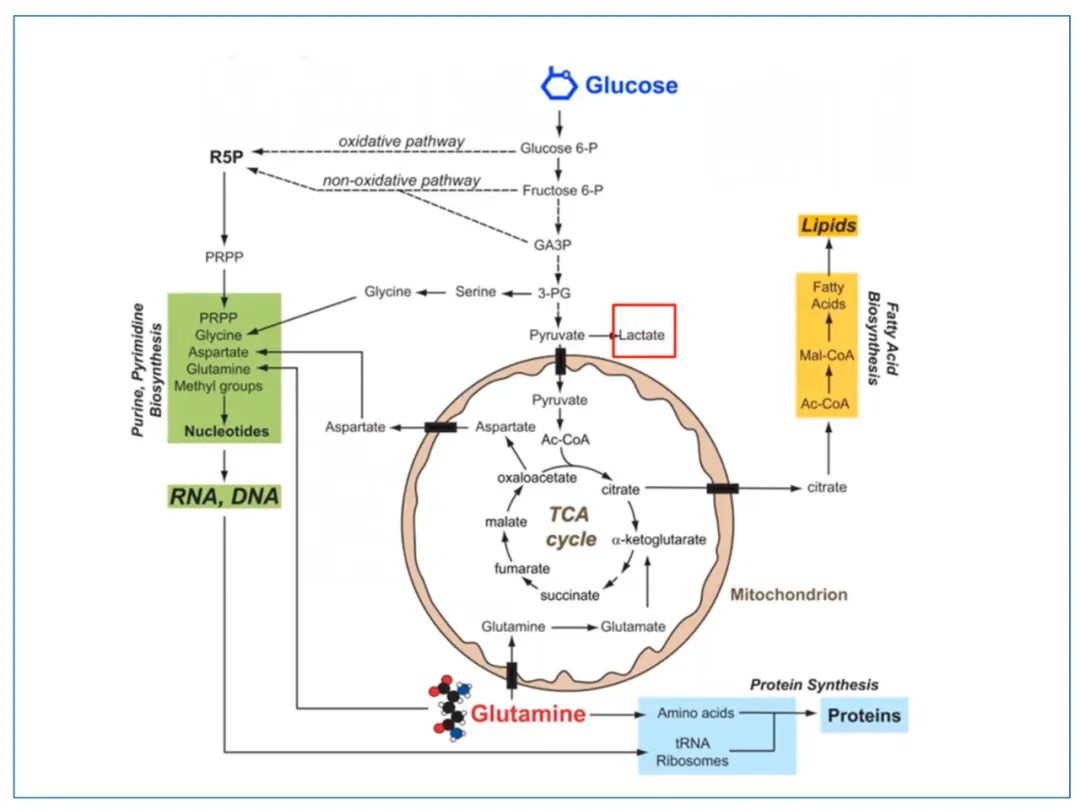
10.谷氨酰胺被运送到线粒体内后,先被转化为谷氨酸,再变成α酮戊二酸(αKG),加入到TCA循环中。TCA循环滚动起来,才能持续地给脂类和核酸合成提供原料。另外,谷氨酰胺还在核酸和蛋白合成中起关键作用。癌细胞为了繁殖,需要源源不断地快速合成大分子。糖酵解和谷氨酰胺摄取都是为了服务这一目的。图源:9

11. 癌细胞具有高度适应性,这也体现在代谢表型上。有些恶性肿瘤细胞表现出高合成代谢率,类似于高度增殖的正常细胞类型。它们摄取大量葡萄糖和谷氨酰胺,并利用这些营养物质驱动TCA循环、OXPHOS、戊糖磷酸途径(pentose phosphate pathway,核苷酸合成)、己糖胺(hexosamine)的合成(参与糖基化分子的合成)、以及氨基酸(蛋白质)和脂质的合成。这些途径共同产生足够水平的细胞成分以支持细胞增殖。有些肿瘤细胞或基质细胞则表现出高分解代谢率。它们牺牲自己,成全大我,通过自噬把自己拆分,为高合成代谢的肿瘤细胞提供乳酸、游离脂肪酸和酮,它们可用于补充 TCA 循环中间体并为 OXPHOS 提供燃料(反向 Warburg 效应)。高合成代谢活性细胞会产生大量的活性氧 (ROS) 。为了维护自身的安全,它们需要产生适当水平的抗氧化剂,包括还原形式的谷胱甘肽(glutathione)。图源:1

12. 这就造成了肿瘤细胞中的代谢异质性。肿瘤组织内的细胞代谢可能会因肿瘤微环境的影响和与血管的距离远近而不同。靠近血液供应的癌细胞优先获取营养和氧气,并通过氧化磷酸化产生 ATP 并上调合成代谢途径,从而支持快速增殖。由这些快速增殖的癌细胞引起的氧化应激会在周围的基质细胞中诱导糖酵解和自噬,从而产生分解代谢物,例如乳酸或酮,它们反过来会被合成代谢型肿瘤细胞吸收,并用于促进线粒体代谢和 ATP 产生。同样,离血管较远的肿瘤细胞接触到的营养较少。它们专注于分解代谢途径,例如自噬,为合成代谢型的肿瘤细胞提供能源和原料。图源:1

13. 肿瘤细胞切换的代谢行为不是孤立的。它和癌症的其它标志相互影响、相互促进。比如,如图7所示,在肿瘤中常见的致癌蛋白激活和肿瘤抑制因子失活都会影响癌细胞的代谢行为。

14. 由于致癌基因或肿瘤抑制因子的突变,酪氨酸激酶受体(TKR)/PI3K信号传导通路是肿瘤中最经常被激活的通路。图源:9

15. TKR或下游的磷脂酰肌醇-3-激酶(PI3K)、Akt的激活使得癌细胞摆脱对生长因子的依赖而自主上调合成代谢活动,包括葡萄糖摄取、糖酵解、脂类合成等。图源:9

16. 肿瘤细胞的异常代谢行为也能帮助它们避免死亡,增加基因组的不稳定性,甚至促进血管生成和癌细胞扩散转移。

17. 在合成代谢型的肿瘤细胞内,由于致癌基因的激活或肿瘤抑制因子的丧失, ROS 产生率增加。ROS对肿瘤细胞来说是一柄双刃剑。适度的ROS水平可帮助肿瘤细胞增殖,并造成更多的基因突变,推动肿瘤细胞的演化。但过高的ROS水平也会杀死肿瘤细胞。因此,在肿瘤的不同阶段,肿瘤细胞都要维持细胞内氧化还原平衡。在增生阶段,肿瘤前体细胞通过诱导 NRF2 依赖性基因的转录来增加抗氧化能力,并在葡萄糖充足的条件下使用戊糖磷酸途径(PPP) 产生 NADPH,从而防止 ROS 积累到自我伤害的水平。当细胞因为血液供应不足而遇到缺氧和低葡萄糖时,ROS 水平会进一步增加。 这时细胞需要 AMPK 和 一碳代谢通路以增强 NADPH 的产生、提高抗氧化能力。当肿瘤突破基底膜,癌细胞逃逸到血液中并传播到远处时, ROS 水平会进一步增加,这时癌细胞需要额外增强抗氧化防御以避免死亡。癌细胞内有过低或过高的 ROS水平都会阻止实体瘤的进展和转移。图源:5

18. 实际上,维持细胞内氧化还原平衡只是肿瘤细胞适应能力的一种。实体瘤在生长过程中会遇到瓶颈,营养和氧气供应不足,比如图中灰色细胞部分。它们会经历代谢压力。因此,肿瘤细胞必须经历一段代谢适应期才能承受这种代谢压力。不能承受压力的细胞通过凋亡而被淘汰。肿瘤的新血管形成是用来缓解这种压力的一种代谢适应策略。图源:24

19. 肿瘤细胞的代谢活动和产物还会影响表观遗传模式,广泛改变基因表达。

20. 新陈代谢和表观遗传学有着千丝万缕的内在联系。组蛋白和DNA的修饰离不开细胞代谢的产物。当葡萄糖进入糖酵解途径时,一小部分分流到己糖胺生物合成途径以产生N-乙酰氨基葡萄糖(GlcNAc)。该 GlcNAc 可成为 OGT酶的底物,将组蛋白 GlcNAc 化。糖酵解通路的活跃程度决定了 NAD+/NADH 比率,这个比率对调节组蛋白脱乙酰酶的活性很重要。包括柠檬酸盐和αKG在内的几种 TCA 循环中间体可以从线粒体中输出。细胞质中的柠檬酸盐转化为乙酰辅酶A,后者用作HAT介导的组蛋白乙酰化的供体。αKG分别被JHDM和TET酶用作组蛋白和 DNA 去甲基化反应的辅助因子。HMT 和 DNMT 的底物是 SAM,由必需氨基酸蛋氨酸合成。另外,低 ATP/AMP 比率可以激活 AMPK,将组蛋白磷酸化。图源:10

21. 代谢产物-表观遗传学轴的重要性在mIDH抑制剂作为抗癌药获批上市中(图5中最后的里程碑)得到验证。正常的异柠檬酸脱氢酶(IDH)催化从异柠檬酸到αKG这一步。在包括AML和胶质瘤等多种癌症中,突变了的IDH将αKG继续转变为正常细胞没有的癌代谢物——2HG。2HG和αKG的结构高度相似,冒着αKG的名义四处招摇撞骗,抑制了所有用αKG作辅助因子的酶。mIDH和2HG最突出的作用就是全面改写表观遗传,使组蛋白和DNA大量甲基化。细胞停留在类似于干细胞阶段,具有快速增殖的能力。mIDH抑制剂可阻止2HG的产生,恢复表观遗传模式。从这一点上来说,Enasidenib (Idhifa) 和 Ivosidenib (Tibsovo)既靶向代谢,又靶向表观遗传。图源:13

22. 现在吸引学术界和工业界最多注意力的是肿瘤细胞的异常代谢对肿瘤免疫微环境的塑造。

23. 前文提到,有氧糖酵解不是肿瘤细胞所独有的。实际上,被激活的T细胞(和髓细胞)也呈现类似的代谢转换:依赖于有氧糖酵解提供ATP, 需求大量葡萄糖和谷氨酰胺,合成更多的大分子以支持细胞增殖……这就引起了肿瘤微环境(TME)中T细胞和肿瘤细胞在代谢上的竞争。图源:4

24. TME是一个由(异质性)肿瘤细胞、各类免疫细胞、基质细胞等构成的生态系统。肿瘤细胞通过掠夺代谢资源和分泌免疫抑制性的代谢产物,扰乱了T细胞的代谢和功能,成功地把客场变成主场,抑制了抗肿瘤免疫活性。因此,一个有效的癌症治疗策略应该包括消除肿瘤细胞代谢带来的负面影响、改善TME的手段。图源:11
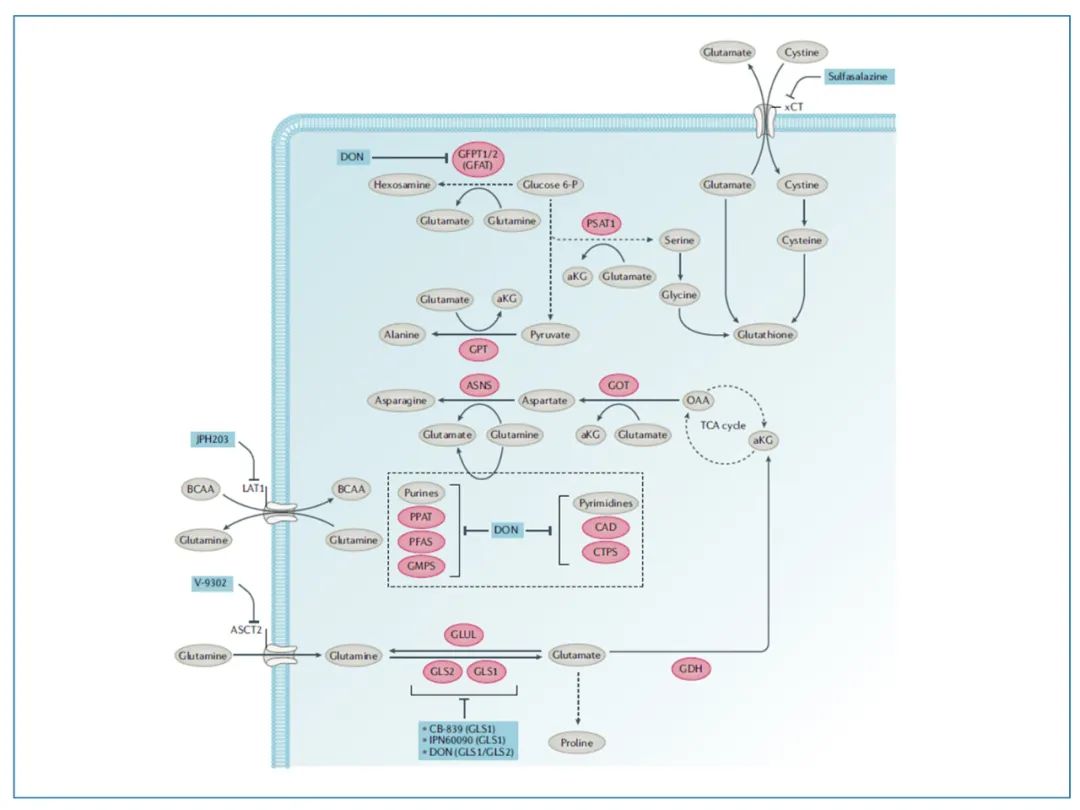
25. 由于肿瘤细胞代谢的复杂性,开发靶向肿瘤代谢的抗癌药物有很大的挑战性。以靶向谷氨酰胺代谢为例,图中红色的元件是谷氨酰胺代谢途径中的代谢酶(潜在靶点)。蓝色元件是现有的小分子抑制剂,能阻断运输蛋白或抑制关键的酶。其中比较有名的是一种谷氨酰胺通路泛阻断剂DON。DON曾在多个针对癌症患者的临床试验中被评估,但终因其毒性太大而搁浅。总体来说,通过抑制谷氨酰胺代谢通路来清除肿瘤细胞目前还有很大争议。图源:16

26. 靶向糖酵解通路也许是另一个治疗癌症的策略,但目前我们还没有得到有力的临床数据来支持这一方向。肿瘤细胞渴望葡萄糖,但红细胞和大脑(和T细胞)也需要糖。阻断糖酵解通路很难避免产生肿瘤外毒性。比如,由于毒性大或疗效低,己糖激酶(HK)抑制剂 2-脱氧葡萄糖的临床开发已被中止。图源:16

27. 这个表中列出了一些靶向癌症代谢的小分子药。其中已批准上市的多是靶向核酸代谢通路的化疗药。表中一个比较有趣的药是抗糖尿病“神药” 二甲双胍(meftormin)。二甲双胍阻断线粒体膜上的电子传输链 (ETC) 复合物 I,抑制氧化磷酸化。二甲双胍在细胞和动物实验中显示了抗癌活性,但临床试验数据好坏参半。目前二甲双胍正在近百个癌症临床试验中被评估。另外,作用机理与其相似的IM156和IACS-010759也已进入临床阶段。
表末的两个化合物indoximod和epacadostat是IDO抑制剂。它们靶向色氨酸分解代谢,降低代谢产物犬尿氨酸(kynurenine),而后者有诱导T细胞衰竭和帮助肿瘤免疫逃逸的功能。也就是说,IDO抑制剂同时靶向代谢和肿瘤免疫。但它们在后期临床试验中尚未取得积极的结果,这再次证明了开发肿瘤代谢药物的挑战性。图源:16

28. 开发肿瘤代谢药物虽然很艰辛,但却是我们征服癌症的必经之路。每个肿瘤的基因突变指纹都可能不同,这导致靶向单一信号通路的药物往往只适用于一小部分癌症患者。但异常的代谢却是大部分肿瘤的共性,因而成为癌症的“阿喀琉斯之踵”。靶向癌症代谢的另一优势是它几乎影响了癌症的所有标志。由于肿瘤细胞的高度异质性和适应性,靶向癌症代谢的药物需要和其它机理的药物联用才能发挥出最佳效果。展望未来,靶向癌症代谢将成为癌症有效治疗策略的一个关键组成部分。图源:adapted from BioRender templates
致谢:
芝加哥大学的陈靖教授对初稿提出了很多宝贵意见和建议。特此表示由衷的感谢。

1. Martinez-Outschoorn, U.E., Peiris-Pages, M., Pestell, R.G., Sotgia, F. and Lisanti, M.P. (2017) Cancer metabolism: a therapeutic perspective. Nat Rev Clin Oncol, 14, 113.
2. Reinfeld, B.I., Madden, M.Z., Wolf, M.M., Chytil, A., Bader, J.E., Patterson, A.R., Sugiura, A., Cohen, A.S., Ali, A., Do, B.T. et al. (2021) Cell-programmed nutrient partitioning in the tumour microenvironment. Nature, 593, 282-288.
3. Ishak Gabra, M.B., Yang, Y., Li, H., Senapati, P., Hanse, E.A., Lowman, X.H., Tran, T.Q., Zhang, L., Doan, L.T., Xu, X. et al. (2020) Dietary glutamine supplementation suppresses epigenetically-activated oncogenic pathways to inhibit melanoma tumour growth. Nature Communications, 11, 3326.
4. Pearce, E.L., Poffenberger, M.C., Chang, C.-H. and Jones, R.G. (2013) Fueling Immunity: Insights into Metabolism and Lymphocyte Function. Science, 342, 1242454.
5. DeBerardinis, R.J. and Chandel, N.S. (2016) Fundamentals of cancer metabolism. Sci Adv, 2, e1600200.
6. Leone, R.D., Zhao, L., Englert, J.M., Sun, I.M., Oh, M.H., Sun, I.H., Arwood, M.L., Bettencourt, I.A., Patel, C.H., Wen, J. et al. (2019) Glutamine blockade induces divergent metabolic programs to overcome tumor immune evasion. Science, 366, 1013-1021.
7. Hanahan, D. (2022) Hallmarks of Cancer: New Dimensions. Cancer Discov, 12, 31-46.
8. Hanahan, D. and Weinberg, R.A. (2011) Hallmarks of cancer: the next generation. Cell, 144, 646-674.
9. Kong, M. (2022), 2022 Cancer Systems Biology Short Course at UCI.
10. Lu, C. and Thompson, C.B. (2012) Metabolic regulation of epigenetics. Cell Metab, 16, 9-17.
11. Leone, R.D. and Powell, J.D. (2020) Metabolism of immune cells in cancer. Nature Reviews Cancer, 20, 516-531.
12. Warburg, O., Wind, F. and Negelein, E. (1927) The Metabolism of Tumors in the Body. J Gen Physiol, 8, 519-530.
13. Golub, D., Iyengar, N., Dogra, S., Wong, T., Bready, D., Tang, K., Modrek, A.S. and Placantonakis, D.G. (2019) Mutant Isocitrate Dehydrogenase Inhibitors as Targeted Cancer Therapeutics. Front Oncol, 9, 417.
14. Eagle, H. (1955) Nutrition Needs of Mammalian Cells in Tissue Culture. Science, 122, 501-504.
15. Hay, N. (2016) Reprogramming glucose metabolism in cancer: can it be exploited for cancer therapy? Nat Rev Cancer, 16, 635-649.
16. Stine, Z.E., Schug, Z.T., Salvino, J.M. and Dang, C.V. (2022) Targeting cancer metabolism in the era of precision oncology. Nature reviews. Drug discovery, 21, 141-162.
17. Farber, S. and Diamond, L.K. (1948) Temporary remissions in acute leukemia in children produced by folic acid antagonist, 4-aminopteroyl-glutamic acid. N Engl J Med, 238, 787-793.
18. Vander Heiden, M.G., Cantley, L.C. and Thompson, C.B. (2009) Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science, 324, 1029-1033.
19. Tran, T.Q., Hanse, E.A., Habowski, A.N., Li, H., Ishak Gabra, M.B., Yang, Y., Lowman, X.H., Ooi, A.M., Liao, S.Y., Edwards, R.A. et al. (2020) α-Ketoglutarate attenuates Wnt signaling and drives differentiation in colorectal cancer. Nature Cancer, 1, 345-358.
20. Powell, J. (2020), SITC 2020.
21. https://ib.bioninja.com.au/higher-level/topic-8-metabolism-cell/untitled/aerobic-overview.html
22. https://metabolicpathways.stanford.edu/
23. www.fishersci.com
24. Jones, R.G. and Thompson, C.B. (2009) Tumor suppressors and cell metabolism: a recipe for cancer growth. Genes & development, 23, 537-548.
25. Cairns, R.A., Harris, I.S. and Mak, T.W. (2011) Regulation of cancer cell metabolism. Nature Reviews Cancer, 11, 85-95.
* 推文用于传递知识,如有版权等疑问,请于本文刊发30日内联系BiG生物创新社。
不感兴趣
看过了
取消

 打赏
打赏
不感兴趣
看过了
取消
精彩评论
相关阅读




中国医院排行榜















 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612



©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:82736610
 京公网安备 11010802020745号
京公网安备 11010802020745号


