
迄今为止,许多研究都集中在人类队列和宏基因组学方法中病毒组的动态。我们现在对与疾病相关的病毒组变化有了一个大致的了解。
导读
全球微生物组与儿科疾病之间的关联已被广泛研究;然而,对微生物组中的病毒成分,即病毒组的研究还不够深入。与病毒组相关的疾病分析通常在技术上具有挑战性,需要对病毒组暗物质进行仔细检查。肠道是病毒颗粒的丰富来源,现在多项研究报告了病毒组与儿科疾病的有趣关联。例如,病毒组研究阐明了与儿童腹泻密切相关的肠道病毒的新谱系,并且儿童IBD病毒组研究开始出现一致的模式。本综述总结了研究病毒组的方法和关于儿童期病毒组性质的最新研究,重点介绍了儿科疾病中肠道病毒组的具体研究。
论文ID
原名:The pediatric virome in health and disease
译名:健康与疾病中的儿科病毒组
期刊:Cell Host & Microbe
IF:21.023
发表时间:2022.5
通讯作者:梁冠翔 & Frederic D. Bushman
通讯作者单位:清华大学医学院;美国宾夕法尼亚大学佩雷尔曼医学院
DOI:10.1016/j.chom.2022.04.006
综述目录
1 前言
2 人体肠道病毒组的组成
3 产前病毒组
4 出生后儿童期病毒组的发育和成熟
5 通过原噬菌体诱导调节病毒组
6 儿科疾病中病毒组的改变
6.1 小儿炎症性肠病
6.2 病因不明的小儿腹泻
6.3营养不良
6.4 自身免疫性疾病
7 结论和展望
主要内容
1 前言
近年来,人们对健康和疾病中的儿科微生物组的了解急剧增加。特别是,已知细菌种群为人类提供营养物质和代谢物,并促进免疫系统的发育,从而影响早期发育和终生健康。偏离健康的群落结构可能与多种儿科疾病有关,如炎症性肠病(IBDs)、肠易激综合征、哮喘、肥胖、自闭症等。
鲜为人知的是,大量病毒(称为病毒组)在生命早期与细菌一起在人体中定植,这些群落的结构也与儿童健康有关。当然,众所周知,致病性病毒会在人体发育的各个阶段引起多种疾病,但在健康婴儿和儿童中存在大量病毒组,偏离健康的群落结构也与疾病有关。
本篇综述首先概述了人类病毒组的组成及其研究方法。接下来,简要总结了人体发育过程中人类肠道病毒组的建立和成熟的研究。然后,我们关注儿科疾病中病毒组的扰动,并审查它们对健康的潜在影响。
2 人体肠道病毒组的组成
病毒组领域日益活跃,并正在面临一系列挑战。例如,在所有病毒基因组中都没有发现可以作为通用标记基因的单一序列。相比之下,细菌和真菌的基因组确实显示出保守区域,允许使用细菌的16S rRNA基因和真菌的rRNA内转录间隔区(ITS)进行分析。因此,病毒组研究依赖于整个病毒群落的宏基因组测序,包括采样、纯化病毒样颗粒(VLPs)、提取病毒核酸、测序和病毒组鸟枪法测序数据的专门分析。
在病毒组序列数据的分析中,病毒“暗物质”的范围是一个显著的挑战。病毒暗物质是指通常在病毒数据中丰富的序列reads,与序列数据库中任何经过充分研究的病毒都不匹配。解决这一问题的一种广泛使用的策略是使用基于机器学习的方法预测组装contig的病毒特征。为此目的开发了许多病毒组分析的算法和方法。最近的研究分析了公开可用的病毒组数据集,使用这些算法对病毒组contigs进行注释,并扩展候选病毒的数据库。
尽管存在这些挑战,但综合研究表明,人类病毒组由几类病毒组成。这些包括感染细菌的噬菌体(以下简称噬菌体)和感染人体细胞的病毒。大约8%的人类基因组包含内源性逆转录病毒的完整病毒序列,病毒也可以在源自食物的肠道标本中发现,例如某些丰富的植物病毒。
能够感染细菌、复制和裂解宿主细胞的噬菌体称为“裂解”病毒。其他能够引发感染,然后将其基因组整合到细菌基因组中的噬菌体被称为温和噬菌体。整合到细菌染色体中的噬菌体基因组称为噬菌体。原噬菌体可以像任何细菌基因一样保持休眠状态并进行DNA复制。在存在诱导信号的情况下,原噬菌体可以恢复裂解生长并裂解宿主细胞。DNA损伤作为一种诱导信号已被广泛研究。促进肠道中噬菌体诱导的信号才刚刚开始研究,并将在后面的综述中讨论。最近,Gregory等人分析了2697个肠道病毒组数据集,得出的结论是,97%以上的序列reads来自噬菌体。噬菌体复制可以遵循几种策略。
噬菌体在新生儿出生后就在肠道中定植,并在整个生命过程中对大部分肠道病毒组做出贡献。真核病毒通常可在3或4个月大的健康婴儿中检测到。影响儿童病毒组动态的因素以及整个病毒群落在早期生命中的潜在作用才刚刚开始被研究。
真核病毒仅占健康人体肠道病毒组的一小部分。常见的病毒类群包括指环病毒科(Anelloviridae)、疱疹病毒科(Herpesviridae)、细小病毒科(Parvoviridae)、多瘤病毒科(Polyomaviridae)、腺病毒科(Adenoviridae)、圆环病毒科(Circoviridae)、杯状病毒科(Caliciviridae)和小核糖核酸病毒科(Picornaviridae)。在肠道中,由于与食物一起食用,病毒也可以出现在粪便样本中,导致发现一些惊人的植物病毒,如帚状病毒科(Virgaviridae)。
病毒可以被脂膜包裹,也可以不被包裹。由于胆汁盐的去污作用和结肠脱水,肠道环境可能会破坏病毒包膜中的脂质双分子层,事实上,肠道中的大多数病毒群都是无包膜的。统计分析表明,病毒粪口传播与无包膜状态有关,大多数噬菌体为无包膜状态。然而,包括SARS-COV-2在内的冠状病毒是个例外,原因尚不清楚。冠状病毒可能在下消化道的肠道细胞中复制,或者,冠状病毒可能具有异常稳定的包膜,在肠道中没有被破坏。
最近的研究将儿科疾病与噬菌体群体结构的改变联系起来,提出了噬菌体在疾病的病理生理学中可能发挥何种功能的问题。噬菌体可能通过调节细菌微生物组或直接与宿主免疫系统相互作用来影响肠道健康。到目前为止,变异病毒组与大多数疾病之间可能存在的因果关系尚不确定。
当然,真核细胞病毒可以是多种疾病的病原体。利用病毒组测序技术寻找病因不明的疾病的可能病毒原因已引起广泛关注。然而,在许多情况下,在病毒样本中检测到的真核病毒与任何疾病都没有联系。例如,在大多数或所有人身上都发现了指环病毒科(Anelloviridae)。它们是小型单链环状DNA病毒,通常可以通过宏基因组方法在生命的许多不同部位进行鉴定。据报道,它们是人类肠道中最早的病毒定植者之一。如果没有上述宏基因组方法对其进行鉴定,这些病毒将是未知的。这些病毒和其他许多病毒在人类健康和疾病中的可能作用尚不确定,其中许多可能是良性共生的。
3 产前病毒组
在健康妊娠期间,子宫内是否存在功能性微生物群仍存在争议,这使得评估可能存在的病毒组变得复杂化。近十年来,随着高通量测序和电子显微镜技术的应用,经典的子宫不育假说受到了挑战。有人提出在人类胎盘、羊水和胎儿中可能存在细菌细胞。最近的一项研究表明,子宫内的微生物组参与了胎儿免疫细胞的初始发育。然而,仔细研究一致表明,各种形式的样本污染可以解释所提出的产前微生物组。此外,无菌剖宫产可产生无菌动物,这一事实为子宫不育提供了强有力的功能证据。
在最近的病毒组研究中,利用下一代测序和荧光显微镜对羊水和胎粪样本的分析不支持出生前存在病毒组。目前的共识是,健康新生儿中不存在产前病毒组,病毒组在出生后迅速获得。
4 出生后儿童期病毒组的发育和成熟
在新生儿时期,肠道中主要的细菌定植者是兼性厌氧菌,如链球菌属(Streptococcus)、葡萄球菌属(Staphylococcus)、肠球菌属(Enterococcus)和埃希氏菌属(Escherichia)。最近的一项研究确定了出生后3年内微生物组成和功能的三个不同成熟期。这包括由母乳、固体食物的引入和其他因素驱动的发育、过渡和稳定阶段。虽然肠道微生物组的一般性质在3岁时变得像成人一样,但在年龄较大的儿童和成人之间仍然可以观察到许多差异。例如,据报道,双歧杆菌(Bifidobacterium)在7-12岁儿童的肠道中富集。儿童中维生素B12合成相关基因的丰度(只存在于微生物中)高于成人。因此,受地理、生活方式、同居、药物等多种因素的调节,婴儿肠道微生物群在大约3年内发育成类似成人的肠道微生物群。
肠道病毒组的成熟仍在评估中(图1)。病毒颗粒总数似乎在出生后迅速达到成人水平。既往研究显示,分娩后病毒颗粒随时间增加,1周内达到~108个/克粪便。1个月后,婴儿肠道内每克粪便含有~109个病毒颗粒,这一水平会持续到成年。最近的一项研究调查了生命早期病毒组的起源,发现许多病毒组序列可以被鉴定为来自同一婴儿肠道细菌基因组中的原噬菌体,这表明来自细菌定植者的诱导原噬菌体参与了婴儿病毒组的大部分。Beller和他的同事们通过密集采样追踪了8名婴儿出生后第一年的肠道病毒组,发现随着年龄的增长,温和噬菌体丰度有显著下降的趋势。另一项较早的研究报告称,母乳传播的几种双歧杆菌中的原噬菌体可以在肠道中诱导。因此,早期婴儿病毒组的组成似乎受溶原诱导支配。
在生命早期,新生儿肠道主要被有尾噬菌体目(Caudovirales)定殖,这在温和噬菌体中很常见。纯裂解噬菌体,如许多微病毒科(Microviridae),通常出现较晚。当肠道细菌种群仍然相对较新且多样性较低时,诸如微病毒科的裂解噬菌体可能很难找到宿主细菌。到2岁时,微病毒科(Microviridae)更普遍,类似于成人肠道病毒组组成。
crAss噬菌体是人类病毒组的另一个重要组成部分。crAss噬菌体通常在50%以上的人类肠道样本中被发现,它们的丰度可以达到肠道病毒群落的90%。crAss噬菌体家族可能是裂解噬菌体,并且与拟杆菌属(Bacteroides)有关。在婴儿出生后的最初几个月内很少能检测到crAss噬菌体,但在较大的婴儿和成人中会变得很普遍,与其宿主的数量相当。
最近的一项研究重新分析了11个公共病毒组数据集,并报告了病毒丰富度的年龄依赖性模式。病毒组在前3年经历了丰富度的下降,在儿童期逐渐增加,在成年期达到顶峰,然后在老年期下降。这一趋势是基于有限的样本(27名0-3岁的健康个体,11名3-18岁的健康个体,93名18-65岁的健康个体,20名65岁以上的健康个体)观察到的,统计检验没有显示婴儿和年龄较大的儿童之间存在显著差异。我们回顾了最近发表的文章,这些研究使用纯化的VLPs分析病毒组,发现对3至18岁儿童的研究相对较少;因此,需要进一步的队列研究来填补这一空白。
最后,最近的证据表明婴儿肠道病毒组可以通过母乳进行调节。母乳含有多种营养物质、微生物、免疫细胞、母体抗体、低聚糖和其他大分子物质。它可以直接抑制真核病毒对婴儿的定植,促进特定细菌物种的生长,从而增加其噬菌体的代表性。另一方面,母乳中可以同时含有真核和原核病毒,这些病毒可以传播到婴儿肠道。几种母体病毒感染通过母乳传播已得到很好的描述,如巨细胞病毒(CMV)、人类免疫缺陷病毒(HIV)和乙型肝炎病毒(HBV)。母乳中报道的其他病毒包括肌病毒科(Myoviridae)、Siphoviridae、短尾病毒科(Podoviridae)和其他真核病毒,并且已经记录了噬菌体从乳房到婴儿肠道的传播。
图1 婴儿肠道病毒组的建立。
上图显示了每克肠道内容物中病毒样颗粒(VLPs)数量随时间的增加。饮食显示在顶部。底部显示了从宏基因组研究推断的相对病毒丰富度曲线。累积的病毒类型由彩色条表示。婴儿个体间在病毒积累的类型和速度方面可能存在很大差异。
5 通过原噬菌体诱导调节病毒组
如上所述,噬菌体基因组可以整合到细菌基因组中形成原噬菌体,诱导是病毒群的主要调控因子。初步计算分析表明,原噬菌体序列可以占细菌基因组的20%。从NCBI数据库(2013年11月更新)检索到的2110个完整的细菌基因组中,近50%包含可识别的假定原噬菌体。最近的一项研究表明,至少有25%的肠道病毒基因组可以被整合到由人类肠道微生物组数据集组装的细菌基因组中,而且超过70%的游离噬菌体有可能是温和噬菌体。从婴儿粪便中分离出的细菌菌株80%以上可诱导释放噬菌体。一项基于培养的研究表明,针对1岁儿童粪便病毒组中大肠杆菌的大多数噬菌体是温和噬菌体,而不是裂解噬菌体。这些研究强调了原噬菌体诱导在肠道病毒组发育中的重要性。
在肠道环境中触发原噬菌体诱导的信号尚不完全清楚。一项研究将溶源性鼠伤寒沙门氏菌(Salmonella typhimurium)暴露于模拟肠道环境中的各种化学物质中,并报道了宿主胆汁盐的诱导作用。一项动物研究表明,肠道炎症通过炎症相关分子(包括活性氧和氮物质)促进沙门氏菌原噬菌体的诱导。Oh及其同事分析了小鼠肠道中益生菌罗伊氏乳杆菌(Lactobacillus reuteri)的原噬菌体诱导。他们报告说,膳食果糖和短链脂肪酸可以提高原噬菌体诱导率。此前的体外研究也强调了其他诱导剂,例如紫外线、抗生素、非甾体抗炎药、群体感应分子、噬菌体共感染以及某些食物和饮料。
原噬菌体也可以自发诱导,尽管诱导率相对较低。最近的一项研究测试了来自婴儿肠道的24种细菌菌株,包括大肠杆菌(Escherichia coli)、克雷伯氏菌(Klebsiella)和肠球菌(Enterococcus)。使用荧光显微镜计数的病毒颗粒由超过30%的菌株自发产生,表明广泛的自发原噬菌体诱导。
因此,在体外和体内都发现了许多原噬菌体诱导物,但对婴儿肠道中原噬菌体诱导信号的研究还很有限。一些诱导剂,如抗生素和非甾体抗炎药,通常在儿童时期使用。对原噬菌体诱导调节儿童肠道微生物群的研究可能会为病毒组调节及其对健康的影响提供相当多的见解。
6 儿科疾病中病毒组的改变
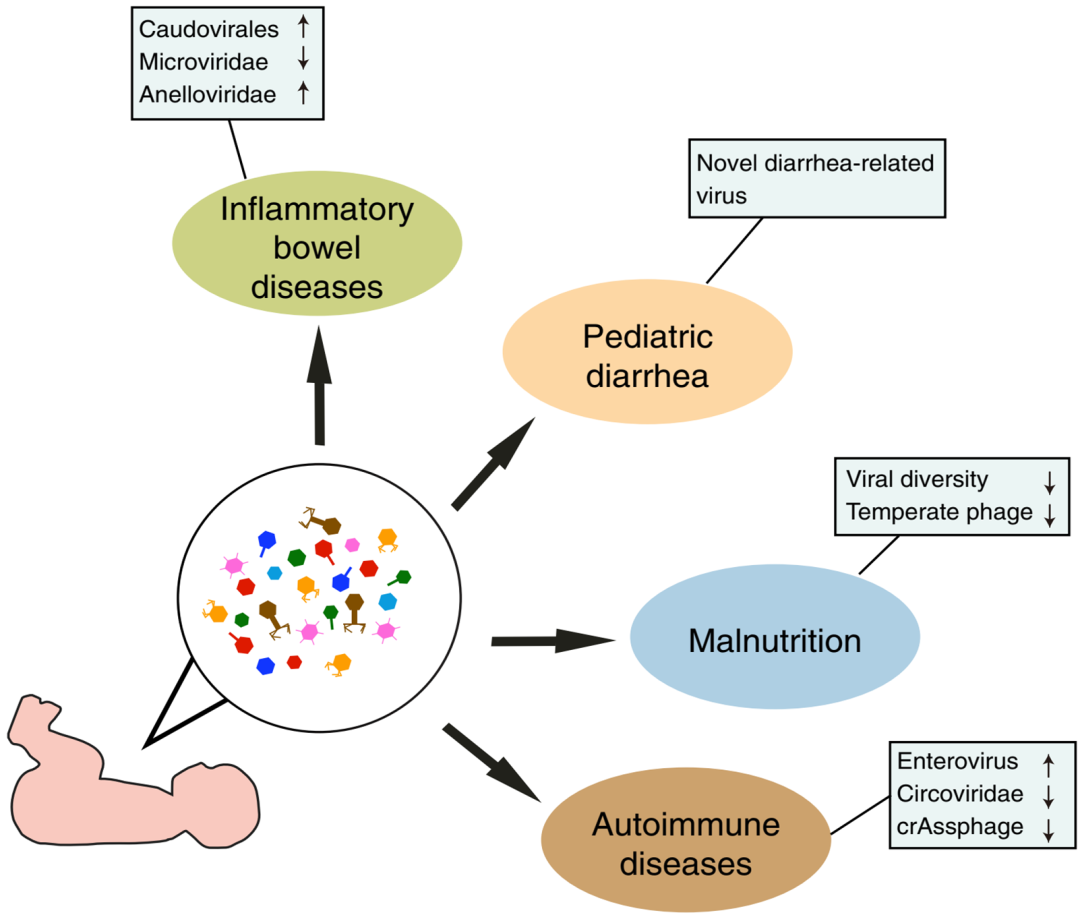
肠道微生物组的改变与多种儿科疾病有关,而儿科微生物群可能与生命后期疾病的发展有关。然而,肠道病毒组的作用在很大程度上是未知的。病毒可以是病原体,通过调节微生物组的其他组成部分促进发病,或与人体免疫系统相互作用,影响对不同疾病的反应。肠道病毒组的动态与多种疾病相关,尽管理解因果关系通常具有挑战性。了解病毒对抗疾病的案例可能是直接可行的,例如,一项研究报告称,粪便病毒组移植可能会减轻小鼠模型中2型糖尿病和肥胖的症状,这就提出了一个问题,即病毒群是否可以直接应用于治疗。在本节中,我们总结了一些关于儿科疾病中人类肠道病毒组的最新发现,包括IBD、病因不明的腹泻、营养不良和自身免疫性疾病(图2;表1)。
图2 与疾病相关的儿童病毒组的改变。
所考虑的疾病显示在椭圆形中,病毒组的变化总结在矩形中。参考文献见正文和表1。
6.1 小儿炎症性肠病
IBDs,包括克罗恩病(CD)和溃疡性结肠炎(UC),是一种慢性肠道炎症性疾病。多项研究报告了IBD病毒组的改变。到目前为止,还没有发现任何病毒病原体。一些感染动物细胞的病毒已被认为是疾病进展的潜在诱因。然而,在不同的研究中发现的病毒有所不同,并没有得出一致的结论。关于小儿IBD,一项研究发现,在儿童CD患者中Caudovirales的丰度增加。另一项研究报告称,在将儿童CD患者与健康个体进行比较时,有尾噬菌体目(Caudovirales)的相对丰度增加,而微病毒科(Microviridae)的相对丰度降低。最近的一项研究分析了一组极早发病的IBD患者(小于6岁),发现了类似的结果,并与成人IBD队列的研究结果相似。这些观察结果的机制仍不清楚。最近的一篇综述文章讨论了可能的解释。一种猜测是肠道炎症触发了原噬菌体的诱导,而一些噬菌体(如Caudovirales)则优先从细菌基因组中诱导。另一种解释是,肠道中噬菌体种群的改变可能直接与宿主免疫系统相互作用,引发或传播肠道炎症。
指环病毒科(Anelloviridae)的富集是IBD中最常报道的真核病毒组特征之一。这也可以在儿科和成人患者中观察到。近年来,IBD中指环病毒科的增加与通常用于治疗IBD的免疫抑制有关,这与其他免疫抑制患者中指环病毒科丰度增加的发现一致。这一观察结果具有潜在的价值,Anelloviridae DNA已被提议作为评估临床免疫抑制疗效的生物标志物。
表1 儿科疾病中肠道病毒组的研究,以及一些主要结论的总结。
6.2 病因不明的小儿腹泻
腹泻是儿童死亡的常见原因,在非工业化国家尤其如此。引起儿童腹泻的病毒病原体包括轮状病毒、诺如病毒、星形病毒、肠道病毒、腺病毒等。轮状病毒是最常见的病毒,2015年造成146 500名5岁以下儿童腹泻死亡。
对腹泻患者的病毒组研究揭示了潜在的新型病毒病原体。例如,一项研究使用病毒组测序技术在布基纳法索腹泻儿童的粪便中发现了一种新的细小病毒,bufavirus。基于PCR的检测显示,在没有轮状病毒的小儿腹泻病例中,bufavirus的流行率为4%。与其他细小病毒相比,该病毒具有高度不同的衣壳蛋白,表明可能存在大量与儿童腹泻有关的bufavirus样病毒,但尚未发现。在另一项分析腹泻儿童和成人真核病毒组的病毒组研究中,这次在荷兰再次发现了bufavirus。在进一步的研究中,Phan等人分析了小儿腹泻患者的粪便病毒组,发现了一组新的小型环状单链DNA病毒基因组,在南美洲和中美洲的患者中流行率从3%到15.5%不等。最近的其他研究在儿童患者中发现了更多与腹泻相关的病毒,如一种新的哺乳动物3型正呼肠孤病毒。
通过功能研究分析这些新型病毒的作用至关重要,与上述病毒相关的病毒也可在健康儿童的肠道中发现,表明亚临床感染的高流行率。因此,了解导致发病的机制和因素将是有价值的。
6.3营养不良
肠道病毒组的改变与儿童营养不良有关。根据世界卫生组织的数据,2019年,5岁以下儿童的死亡中约有45%是营养不良造成的。与健康个体相比,营养不良儿童的肠道微生物群发生了改变。以微生物群为导向的食品已被设计用于治疗营养不良,产生了可喜的结果,强调了微生物群的重要性。
最近的一项研究分析了8对健康生长的双胞胎和12对严重急性营养不良(SAM)的双胞胎的肠道病毒组。双胞胎队列研究很有价值,因为他们可以潜在地将与遗传背景相关的表型与定植的随机性区分开来。该研究报道了SAM不协调双胞胎对噬菌体种群和真核病毒(Anelloviridae和Circoviridae)的可区分特征。在不一致的双胞胎中,可以发现两个成员的病毒组结构都被破坏,这表明这些表面上健康的孩子可能有更高的发展为营养不良的风险。
发育不良儿童的病毒组(营养不良的一种亚型)也已被研究。Desai等人鉴定了3种不同生长速度下差异丰富的Caudovirales物种。他们还报道称,在生长良好的儿童中观察到的细菌和噬菌体多样性之间的正相关关系在生长不良的儿童中被破坏了。另一项研究报告称,与未发育不良的对照组相比,发育不良儿童体内的噬菌体多样性较低,温和噬菌体数量较少。体外交叉感染实验表明,纯化的噬菌体可以感染细菌群落并调节其丰度和组成。上述两项研究都表明,发育不良儿童肠道内噬菌体种群的改变可能通过调节细菌群落来影响宿主,而细菌种群的变化可能会影响宿主代谢或促进肠道炎症,从而导致生长受损。这些观察结果背后的确切机制仍有待确定。
6.4 自身免疫性疾病
1型糖尿病是一种自身免疫性疾病,通常发生在儿童期和青年时期,其中产生胰岛素的β细胞受到自身免疫系统的攻击。1型糖尿病的病因仍然未知,而病毒被认为是潜在的触发因素。胰岛自身免疫描述了1型糖尿病发病前血清中出现的胰岛自身抗体,这是一种可能导致1型糖尿病的自身免疫过程。肠道病毒与该疾病有关,因为它是从儿童1型糖尿病患者的胰腺中分离出来的一个病毒家族,并且观察到肠道病毒在患1型糖尿病的兄弟姐妹中比在非糖尿病兄弟姐妹中更流行。其他病毒,如轮状病毒、腮腺炎病毒、风疹病毒和巨细胞病毒,也与1型糖尿病有关。
然而,不同研究的结果不同。一项初步研究调查了19名2岁以下胰岛自身免疫儿童的粪便病毒组,并没有发现患病个体和健康对照之间存在任何显著差异。Zhao等人追踪了1型糖尿病高风险儿童从出生到2-3岁的肠道病毒组,发现1型糖尿病高风险儿童中Circoviridae相关的contigs减少。他们还报道了与疾病相关的噬菌体contigs,它们与细菌丰度呈正相关或负相关。另一项研究记录了胰岛自身免疫儿童中拟杆菌的减少以及crAss噬菌体和Bacteroides dorei之间的正相关关系。最近的一项研究对粪便和血浆样本进行了病毒组分析,发现与对照组相比,患病儿童的肠道病毒丰度更高。这与之前的非病毒组测序研究相似,表明肠病毒感染通常先于1型糖尿病的发病。最近的另一项研究对383名胰岛自身免疫儿童和112名出现1型糖尿病症状的儿童的粪便样本进行了真核病毒组分析,并表明长期肠道病毒感染与胰岛自身免疫有关,但与1型糖尿病无关。
总之,虽然肠病毒感染在1型糖尿病中很常见,但目前尚不清楚肠道病毒是否为致病因子。肠道病毒频繁感染还与其他自身免疫性疾病有关(如乳糜泻),这表明肠道病毒感染并不是特异性的,甚至可能是自身免疫性疾病的结果。已经报道了肠道噬菌体种群的改变,这增加了噬菌体可能导致1型糖尿病发展的可能性。
7 结论和展望
自2008年Breitbart等人发表第一篇论文以来,儿科病毒组研究取得了显著进展,其中Breitbart等人使用宏基因组测序技术描述了新生儿的粪便病毒群落。如今,我们对人类病毒组在出生后的初始播种、头3年病毒组成的动态以及噬菌体-细菌相互作用已经有了初步的了解。但还需要更多的队列研究来完全确定病毒组进入成年的成熟过程。有报道称,原噬菌体诱导可通过多种方式调节细菌群落,新出现的数据表明,原噬菌体诱导可能是儿童肠道微生物群的重要调节因子。母乳在塑造婴儿病毒组方面起着至关重要的作用,而饮食通常是成人和儿童病毒组的调节剂。更详细地了解肠道中存在的不同诱导信号以及正在进行的噬菌体诱导如何调节微生物群落结构和健康结果将是非常有意义的。
在探索肠道病毒组和儿科疾病之间的关系方面已经取得了进展,但仍有许多需要研究的地方。表1总结了儿童肠道病毒组和儿科疾病之间关联的显著例子。病毒组测序为发现原因不明的儿科疾病的候选病毒病原体提供了强有力的方法。迄今为止,许多研究都集中在人类队列和宏基因组学方法中病毒组的动态。我们现在对与疾病相关的病毒组变化有了一个大致的了解。未来的方向可以侧重于探索因果效应,例如,在精心设计的动物模型。由于确定的噬菌体群落与疾病呈负相关,可以在人类干预研究中将它们作为先进的噬菌体疗法进行测试。因此,正在进行的研究将有可能产生创新的新疗法。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您


































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




