
炎症小体促进炎症相关疾病,包括癌症,并有助于放疗引起的组织损伤。然而,炎症小体在放疗诱导的抗肿瘤效应中的作用尚不清楚。
炎症小体促进炎症相关疾病,包括癌症,并有助于放疗引起的组织损伤。然而,炎症小体在放疗诱导的抗肿瘤效应中的作用尚不清楚。2021年5月,Chuanhui Han等人在《Sci Immunol》杂志上发表了一篇题为《The AIM2 and NLRP3 inflammasomes trigger IL-1–mediated antitumor effects during radiation》的文章,研究了放疗对几种炎症小体缺陷小鼠的抗肿瘤作用,以明确炎症小体通路中哪些分子参与了对放疗的抵抗。现介绍如下:
背景
核苷酸结合和寡聚化结构域(NOD)样受体(NLR)家族中有几种胞浆模式识别受体(PRRs)参与炎症小体的形成和激活,包括NLRP1,NLRP3,NLRP6和NLRC4。其中NLRP3是最具特征的NLR,可以识别广泛的PAMPs和DAMPs,包括DNA、5'-三磷酸腺苷和活性氧(ROS)。黑素瘤2(AIM2)中缺失的胞浆DNA传感器是一个形成PRR的非NLR炎症小体,它也参与炎症小体的激活。NLRP3或AIM2通过其特异性配体激活后,通过含有C端caspase募集结构域(ASC)的适配体蛋白凋亡相关斑点样蛋白与caspase-1 ( CASP1 )相互作用,形成大型多分子炎症小体复合物。这种相互作用激活CASP1,使IL-1β和IL-18裂解,产生其成熟形式。IL-1β发挥多种生物学功能,包括激活淋巴细胞和募集中性粒细胞到局部组织。IL-1β与肌层细胞或T细胞上的IL-1R1结合后,通过激活核因子κB、丝裂原活化蛋白激酶和激活蛋白1促进趋化因子和促炎细胞因子的分泌。趋化因子和细胞因子的分泌在感染部位或肿瘤组织中建立了炎症微环境。
与许多急性感染引起的促炎免疫反应不同,肿瘤促进慢性炎症建立免疫抑制性肿瘤微环境(TME)。值得注意的是,炎症小体在肿瘤引起的免疫抑制和肿瘤发生中发挥作用,特别是在炎症引起的癌症中。此外,新的证据表明,阻断炎症小体介导的IL-1信号可能有助于控制肿瘤生长。Casp1−/−小鼠或阻断IL-1信号时,肿瘤生长较慢。一些化疗药物,如吉西他滨和5-氟尿嘧啶,激活NLRP3炎症小体,导致IL-1β的分泌。用IL-1R拮抗剂阻断IL-1R信号可以增加吉西他滨和5-氟尿嘧啶的抗肿瘤作用,并减少肿瘤的发展和转移。阻断IL-1R信号作为癌症治疗手段在临床试验中被广泛研究。
放疗( RT )作为癌症治疗手段被广泛应用。放疗疗效主要取决于直接杀伤肿瘤细胞和随后诱导的免疫反应,这些反应目前还不清楚。T细胞介导的抗肿瘤免疫有助于局部消融RT治疗后的肿瘤抑制。肿瘤源性双链DNA (dsDNA)可通过干扰素基因(STING)通路的cGMP (鸟嘌呤-5 '-单磷酸)-AMP(5’单磷酸腺苷)合成酶(cGAS)-刺激剂感知,导致产生I型干扰素(IFNs),其在交叉激活、T细胞活化、肿瘤的后续消退中起核心作用。除了cGAS, DNA可以被其他几种PRRs感知,但这些PRRs及其相关免疫通路在RT期间的抗肿瘤免疫中的确切作用尚不清楚。AIM2和NLRP3有助于放疗诱导的细胞死亡和组织损伤。病毒感染巨噬细胞可诱导AIM2炎症小体活化,限制了cGAS-STING通路。这表明炎症小体可能抑制RT介导的抗肿瘤免疫反应。然而,在这里,作者发现Casp1−/−小鼠的肿瘤生长比对照小鼠慢,并且对RT抵抗。具体来说,放疗诱导AIM2和NLRP3炎症小体的活化,并协同诱导巨噬细胞产生IL-1,进而激活DC。这种DC激活导致了更强的T细胞免疫应答和稳健的抗肿瘤免疫。
结果
CASP1信号增强 了放疗 的抗肿瘤作用
考虑到炎症小体在限制cGAS-STING激活中的作用,作者推测小鼠体内CASP1的缺失会通过刺激cGAS-STING通路的激活来限制肿瘤生长并增强RT。作者观察到,MC38移植瘤在Casp1−/−小鼠中生长比野生型(WT)小鼠慢(图1A)。为了验证CASP1缺失是否对RT诱导的肿瘤抑制有好处,作者研究了MC38肿瘤在WT或Casp1−/−小鼠中照射后的生长情况。出乎意料的是,Casp1−/−小鼠肿瘤对RT的反应不如WT小鼠(图1B)。由于Casp1−/−小鼠体内肿瘤生长较慢,希望排除RT的抗肿瘤作用不是由于WT和Casp1−/−小鼠之间肿瘤生长动力学的改变。因此,在Casp1−/−小鼠体内接种了两倍于WT小鼠的肿瘤细胞,这导致在放疗处理时肿瘤大小相似。与最初的观察类似,放疗对Casp1−/−小鼠移植瘤生长的控制并不有效(图1C),还用B16-卵清蛋白(OVA)肿瘤评价了CASP1在RT诱导肿瘤抑制中的作用。未处理的B16-OVA肿瘤在WT和Casp1−/−小鼠中生长相似;然而,放疗并不能有效控制Casp1−/−小鼠的B16-OVA肿瘤(图1D)。这些数据提示,CASP1在小鼠体内的表达在放疗诱导的抗肿瘤活性中起关键作用。
图1 WT和Casp1 -/- 小鼠辐照肿瘤的生长情况
放疗引发 AIM2和NLRP3炎症小体 的激活
放疗引起DNA损伤和细胞死亡,使胞内和胞外dsDNA增加。由于AIM2在感知dsDNA后形成炎症小体,作者想知道RT是否激活了TME中的AIM2炎症小体。为了解决这一问题,将MC38肿瘤细胞移植到Aim2-/-小鼠体内,并对肿瘤进行放疗治疗。与Casp1-/-小鼠的肿瘤不同,Aim2-/-小鼠的MC38肿瘤对放疗仍然敏感(图2A)。放疗处理增加ROS的产生,释放线粒体DNA到胞浆中,诱导Ca2+和K+内流,从而激活NLRP3炎症小体。因此,作者研究了宿主NLRP3在RT过程中是否发挥抑瘤作用,发现Nlrp3-/-小鼠的MC38肿瘤对放疗的反应与WT和Aim2-/-小鼠相似(图2A)。
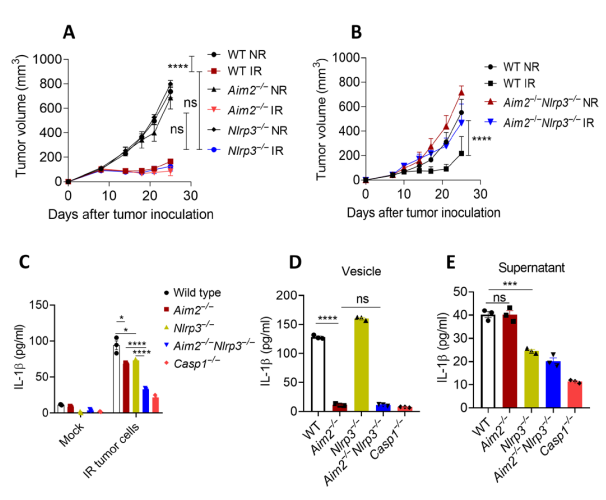
在骨髓移植模型中,放疗同时激活AIM2和NLRP3炎症小体。在上述结果的基础上,作者提出在体外系统中鉴定免疫细胞中AIM2和NLRP3炎症小体激活对辐照肿瘤释放的DAMPs的相对贡献。因此,作者将辐照的MC38肿瘤细胞与WT、Casp1−/−或Aim2−/−巨噬细胞共培养。通过酶联免疫吸附法(ELISA)检测发现,照射后的肿瘤细胞可诱导WT产生IL-1β,而不是Casp1−/−巨噬细胞,说明照射后肿瘤源性介质激活了炎症小体。与WT巨噬细胞相比,Aim2−/−巨噬细胞部分减少了IL-1β的产生。作者还在WT和Aim2−/−巨噬细胞中使用小分子抑制剂MCC950抑制Nlrp3炎症小体。MCC950进一步减少了WT和Aim2−/−巨噬细胞IL-1β的产生。这些数据表明,AIM2和NLRP3在放疗诱导的炎症小体活化和IL-1β的产生中起互补作用。
为了验证AIM2和NLRP3在IL-1β产生中起互补作用的假设,作者制备了Aim2−/−Nlrp3−/−小鼠,其Aim2和Nlrp3基因表达均缺失,Western blotting证实了这一假设。将MC38肿瘤移植到Aim2−/−Nlrp3−/−小鼠中。在Aim2−/−Nlrp3−/−小鼠中,肿瘤对放疗具有抵抗性,这表明当Aim2和Nlrp3均失活时,放疗诱导的肿瘤生长抑制受到了损害(图2B)。为证实Aim2和Nlrp3双重缺失对放疗诱导的CASP1活化的影响,作者将WT、Casp1−/−、Nlrp3−/−、Aim2−/−和Aim2−/−Nlrp3−/−小鼠巨噬细胞与放疗处理的MC38细胞共培养,并通过ELISA法检测成熟IL-1β的水平。与Aim2−/−Nlrp3−/−小鼠的放疗反应一致,双敲除小鼠IL-1β产生减少,而Aim2−/−或Nlrp3−/−巨噬细胞仅部分减少IL-1β的产生(图2C)。因此,AIM2和NLRP3均协同调节RT介导的抗肿瘤活性。
接下来,作者研究了辐照肿瘤细胞刺激巨噬细胞中AIM2和NLRP3炎症小体活化的机制。由于细胞分子和细胞器可以直接或通过细胞外囊泡(EVs)分泌,所以作者想要检测EV或非EV组分是否激活炎症小体。因此,收集辐照MC38细胞的培养液,从培养上清中纯化EVs,并分离出无EV的上清。然后将WT或炎性体缺陷小鼠巨噬细胞与EV或无EV上清液共同孵育。辐照MC38肿瘤细胞的EVs显著增加了WT细胞IL-1β的产生。Casp1−/−和Aim2−/−Nlrp3−/−巨噬细胞中未见IL-1β升高。这些数据提示,辐照肿瘤细胞源性EVs可激活AIM2和/或NLRP3炎症小体。Aim2而非Nlrp3参与了EV诱导的IL-1β的产生 (图2D),表明辐照的肿瘤细胞源性EV激活了巨噬细胞中的Aim2炎症小体。与此相反,无EV上清以Nlrp3依赖的方式诱导IL-1β的产生(图2E)。这些数据提示,辐照肿瘤通过EV和非EV组分同时触发了髓系细胞中AIM2和NLRP3炎症小体的激活。
图2 AIM2和NLRP3炎症小体对RT的作用
RT诱导的抗肿瘤免疫 需要 宿主 体内的 IL-1信号 通路
作者观察到,辐照肿瘤可触发巨噬细胞内AIM2和NLRP3炎症小体活化,这对RT的抗肿瘤作用至关重要。作者感兴趣的是探索炎症小体介导RT抗肿瘤活性的内在机制。炎症小体导致两种关键炎性细胞因子IL-18和IL-1β裂解,因此,作者着重测定IL-18和IL-1β在RT过程中的抗肿瘤作用。对Il18r1−/−、Il18−/−或Il1r1−/−小鼠的MC38肿瘤进行照射,并监测肿瘤生长,发现Il18r1−/−和Il18−/−小鼠的肿瘤对RT敏感(图3A),而Il1r1−/−小鼠的放疗对辐射具有抗性(图3B)。这些观察表明IL-1信号而不是IL-18信号参与了RT的抗肿瘤活性。
由于IL1R1同时识别IL-1β和IL-1α,作者研究了哪种细胞因子在放疗介导的抗肿瘤免疫应答中发挥关键作用。为了解决这个问题,用重组抗体阻断了IL-1β在WT小鼠体内的表达,并评估了放疗诱导的肿瘤消退。IL-1β阻断不影响放疗疗效。作者推测这种影响可能是由IL-18和IL-1β的互补作用引起的,因此,在Il18r1−/−小鼠中检测了阻断IL-1β后RT的抗肿瘤活性。出乎意料的是,IL-1β阻断后,Il18r1−/−小鼠中MC38肿瘤的放疗敏感性未发生改变 (图3C)。这些观察出乎意料,但指出了其他IL1R1上游细胞因子可能参与增强放疗的治疗效果。AIM2和NLRP3炎症小体也调节IL-1α的产生,IL-1α也通过IL1R1发出信号,因此,作者研究了IL-1β和IL-1α在放疗介导的肿瘤生长抑制中的协同作用,发现阻断IL-1β或IL-1α均不影响放疗的疗效,但同时阻断IL-1β和IL-1α可降低肿瘤的放射敏感性(图3D)。为了探讨CASP1是否也对IL-1α的产生至关重要,作者随后检测了共培养体系和肿瘤组织中IL-1α的蛋白水平。发现Casp1−/−BM源性巨噬细胞(BMDM)与辐照MC38肿瘤细胞共培养时产生的IL-1α比WT BMDM少;此外,荷瘤Casp1−/−小鼠经放疗后血清IL-1α水平也低于WT小鼠。这些结果表明IL-1信号在放疗诱导的肿瘤生长抑制中发挥重要作用。
图3 探讨IL-1通路对RT的影响
IL-1通过刺激DCs触发抗肿瘤T细胞反应
接下来,作者研究了哪些免疫细胞类型参与了CASP1/IL1R1介导的RT抗肿瘤作用。由于IL1R1信号在T细胞活化中起关键作用,首先利用Cd4-Cre;Il1r1fl/fl小鼠(T细胞中特异性丢失Il1r1)评估了T细胞固有IL-1信号在放疗诱导抗肿瘤免疫中的作用,出乎意料的是,T细胞中IL-1信号的丢失仅略微降低了MC38肿瘤对放疗的敏感性(图4A),这表明IL-1主要靶向T细胞以外的细胞,以促进放疗的抗肿瘤作用。巨噬细胞在早期发挥RT的抗肿瘤作用中起重要作用;然而,它们还涉及建立一个免疫抑制TME来支持RT晚期的肿瘤生长。作者生成了巨噬细胞中Il1r1缺失的Lyz2-Cre;Il1r1fl/fl小鼠。与Il1r1fl/fl小鼠一样,Lyz2-Cre;Il1r1fl/fl小鼠对放射治疗也有反应(图4,B和C)。这表明IL-1并不靶向巨噬细胞以利于放射治疗。主要的抗原呈递细胞,DCs,在启动肿瘤特异性T细胞反应和RT诱导的抗肿瘤免疫中起关键作用。因此,作者生成了DCs中特异性缺乏IL1R1信号的zDC-Cre;Il1r1fl/fl小鼠,发现其对RT的反应较差,提示DC中放疗诱导的Casp1/IL-1信号在放疗诱导的抗肿瘤免疫应答中起一定作用(图4D)。
图4 IL-1靶向DC增强放疗介导的抗肿瘤免疫
放疗诱导肿瘤生长抑制需要CD8+T细胞。DCs可以通过交叉启动激活肿瘤特异性CD8+T细胞,DC特异性IL-1信号在放疗治疗中的重要作用使作者怀疑IL-1是否靶向DCs调节CD8+T细胞的功能。放疗时肿瘤内的CD8+T细胞对放疗具有耐药性,放疗时阻断CD8+T细胞的浸润并不影响放疗的治疗效果。这说明,放疗前已有的CD8+T细胞,而不是新浸润的CD8+T细胞,可能是放疗诱导抗肿瘤免疫的主要效应分子。因此,作者推测DCs中IL-1信号是启动受照射肿瘤特异性瘤内CD8+T细胞所必需的。为探讨这种可能性,从脾细胞中分离出OVA肽SIINFEKL(OT-1)特异性CD8+T细胞,并对其进行照射,以模拟照射后的瘤内CD8+T细胞;然后,OVA预处理的DCs与照射后的OT1 CD8+T细胞共培养。与WT DC (图5,A和B)相比,Il1r1−/−DCs显著减少CD8+T细胞产生IFN-γ和IL-2。IL-1β处理进一步增加了与WT共培养的CD8+T细胞中IFN-γ和IL-2的产量,而不是Il1r1−/−DCs(图5,A和B)。IL-1β处理也增加了与WT共培养的CD8+T细胞的存活率,但Il1r1−/−DCs没有(图5C)。为证实这一观察,作者还调查了放疗后原有的肿瘤CD8+T细胞的存活情况。IL1R1信号不影响巨噬细胞或DCs的存活;然而,与WT小鼠相比,Illr1−/−小鼠的T细胞较少(图5,D和E)。这些数据共同表明,放疗激发了AIM2和NLRP3炎症小体的活化,并以CASP1依赖的方式促进IL-1的产生。通过促进DC的交叉启动功能,IL-1进一步激活了原有的CD8+T细胞。
图5 DCs中IL-1信号有助于RT后原有CD8+T细胞的存活
IL-1β 可以 克服肿瘤的 放疗 抵抗
考虑到DCs中IL-1信号通路在RT抗肿瘤活性中的重要作用,作者想检测局部给予IL-1β是否可以逆转Casp1−/−和Aim2−/−Nlrp3−/−小鼠肿瘤的放疗抗性。将MC38细胞接种到Casp1−/−小鼠中,然后对肿瘤进行放疗、IL-1β、或放疗和IL-1β联合治疗。移植的MC38肿瘤对放疗或IL-1β治疗均耐药(图6A)。放疗和IL-1β联合治疗克服了Casp1−/−小鼠肿瘤的放疗抗性(图6A)。同样,给予IL-1β也能使Aim2−/−Nlrp3−/−小鼠的MC38肿瘤对RT敏感(图6B),为了验证IL-1β对WT小鼠RT的影响,作者还在WT小鼠体内接种MC38,对肿瘤进行放疗和/或IL-1β处理。同样,IL-1β与放疗协同控制WT小鼠的MC38肿瘤生长(图6C)。然而,Illr1−/−小鼠的肿瘤仍然对放疗和IL-1β单独或联合治疗耐药(图6D),这表明通过靶向宿主免疫细胞的IL-1信号而发挥IL-1β的作用。为了进一步验证IL-1信号在DC联合治疗中的作用,作者在zDC-Cre;Il1r1fl/fl小鼠中接种了MC38肿瘤细胞,与Il1r1−/−小鼠的结果一致,zDC-Cre;Il1r1fl/fl小鼠的肿瘤对放疗和IL-1β的联合治疗耐药(图6E)。这些发现提示,给予IL-1β靶向DCs克服AIM1/NLRP3或CASP1缺陷小鼠的放疗抗性。
图6 RT后IL-1β的抗肿瘤作用
总之,作者证明了AIM2和NLRP3炎症小体在放疗诱导的抗肿瘤免疫中的重要作用。通过AIM2和NLRP3炎症小体放疗诱导CASP1活化后,IL-1β的生成增加,刺激DCs激活辐照肿瘤中的CD8+T细胞,并诱导抗肿瘤免疫。
讨论
在此,作者证明AIM2和NLRP3炎症小体协同增强RT诱导的抗肿瘤免疫。与Casp1−/−小鼠肿瘤的放射抗性不同,Aim2−/−或Nlrp3−/−小鼠的肿瘤对放疗有反应。然而,AIM2和NLRP3的缺失降低了RT的治疗效果。机制上,辐照肿瘤源性EVs诱导AIM2炎症小体活化,而无EV上清主要激活NLRP3炎症小体。RT后AIM2和NLRP3炎症小体的激活导致IL-1β成熟,增强DCs的交叉启动活性,从而激发受照射肿瘤中CD8+T细胞的功能。额外的IL-1β治疗有助于克服Casp1−/−和Aim2−/−Nlrp3−/−小鼠肿瘤的放射抗性。同时,放疗通过AIM2和NLRP3炎症小体激活CASP1,通过增强DCs中的IL-1信号来增强抗肿瘤免疫反应。
放疗的抗肿瘤作用依赖于直接杀伤肿瘤细胞和放疗的免疫刺激作用。T细胞介导的抗肿瘤免疫有助于局部消融RT的肿瘤抑制。肿瘤来源的dsDNA可通过DNA传感通路被感知,导致I型IFNs的产生,在DC交叉启动、T细胞激活和随后的肿瘤消退中起核心作用。炎症小体是cGAS-STING通路的负调节因子,因此,由于cGAS-STING通路的激活程度较高,CASP1缺失有望增强RT活性。相反,作者观察到RT在Casp1−/−和Il1r1−/−小鼠中抑制肿瘤生长的效果较差。这表明CASP1-IL-1轴以一种不依赖于cGAS-IFN的方式激发RT介导的抗肿瘤免疫。
NLRP3和AIM2炎症小体与IFNs通路的调控关系复杂。在cGMP-AMP或IFN刺激后,AIM2、NLRP3、CASP1和IL-1β在脂多糖诱导的BMDMs中上调。cGAS-STING也能以不依赖于I型IFN的方式激活NLRP3炎症小体。相反,I型IFN抑制NLRP1和NLRP3炎症小体的活化以及pro-IL-1α和pro-IL-1β的转录。目前,CASP1是否也通过抑制cGAS-STING通路影响RT还有待确定。本研究发现,EVs激活巨噬细胞中的AIM2炎症小体。辐照的肿瘤源性EVs也可能启动cGAS- STING通路,因为cGAS与AIM2一样也是胞质dsDNA传感器。考虑到炎症小体对cGAS- STING通路的抑制作用,AIM2和cGAS的激活可能以细胞类型特异性或以时间的方式发生,或两者兼而有之。
除cGAS- STING介导的I型IFN信号外,作者还发现AIM2/ NLRP3- CASP1介导的DCs IL-1信号参与了RT的放疗抗肿瘤作用。I型IFNs信号在DCs的交叉启动功能中起基础性作用。DCs中I型IFN信号的缺失会抑制CD8+T细胞的活化。作者观察到,DCs中IL-1信号的丢失减少了辐照CD8+T细胞的交叉启动。这就提出了另一个关键问题,即与放疗后新浸润的CD8+T细胞相比,肿瘤内的CD8+T细胞在放疗后如何存活并发挥作用。预先存在的CD8+T细胞在RT诱导的抗肿瘤免疫中起关键作用,具有类似组织驻留CD8+T细胞的基因表达模式。本研究在RT背景下揭示了IL-1信号在DCs中的作用,然而,DC中IL-1信号调节辐照CD8+T细胞功能的确切机制尚不清楚。与Il1r1−/−DCs相比,WT DCs显著增加了CD8+T细胞IL-2的产生,这可能也有助于CD8+T细胞的活化和存活。此外,本研究表明,DCs对辐照肿瘤中CD8+T细胞的存活至关重要。大多数辐照CD8+T细胞在体外培养2~3天后死亡;然而,在与DCs共培养时,约15~30%的T细胞仍然存活。IL-1等因子如何调控肿瘤原有的CD8+T细胞功能的分子机制尚待确定。
“论肿道麻”述评
炎症小体是一种胞浆多蛋白复合物,是响应病原体和危险相关分子模式(PAMPs和DAMPs)而形成的,在宿主防御感染和肿瘤发生中起关键作用。作为炎症小体的主要产物,IL-1β首先被鉴定为T细胞活化因子,并首次被证实在肉瘤、黑色素瘤和腺癌模型中具有抗肿瘤作用。然而,临床试验并没有证实IL-1治疗的有效性,而且它有剂量相关的毒性。该研究结果提示,放疗诱导的肿瘤控制需要DCs中的急性IL-1信号;局部给予IL-1β与放疗协同克服AIM2/NLRP3-CASP1轴的缺陷。虽然全身性给予IL-1β能有效触发免疫反应,但具有严重的毒性。局部或靶向给予IL-1β的策略与放疗联合使用时可能有协同效应,且毒性比全身性给予低。
虽然炎症小体有助于肿瘤免疫微环境,但炎症小体在放疗诱导的抗肿瘤作用中的作用尚不清楚。在这里,作者发现放疗诱导的AIM2和NLRP3炎症小体的激活激发了DC细胞上的IL-1信号,增强了T细胞免疫。该研究可能为增强CASP1-IL-1β信号用于癌症免疫治疗开启更多的研究。
编译:罗桓宇;述评:周国霞
审核:张军,缪长虹

参考文献: The AIM2 and NLRP3 inflammasomes trigger IL-1–mediated antitumor effects during radiation . Han C, Godfrey V, Liu Z , et al. Sci Immunol . 2021 May ; 6 ( 59 ). doi: 10.1126/sciimmunol.abc6998 . Epub 202 1 May 7.
( 向上滑动查看内容 )
声明:古麻今醉公众号为舒医汇旗下,古麻今醉公众号所发表内容之知识产权为舒医汇及主办方、原作者等相关权利人所有。未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。欢迎转发、分享。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您







































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




