

基于宏基因组组装基因组和单细胞RNA测序奶牛瘤胃纤维利用研究



导读
背景:奶牛利用人类不可食用的低价值植物生物质来生产牛奶,这是一种营养丰富、蛋白质含量高的低成本产品。这一过程很大程度上依赖于瘤胃微生物发酵木质纤维素和纤维素来产生挥发性脂肪酸(VFAs)。VFAs在不同细胞类型的介导下,被瘤胃分层鳞状上皮吸收并部分代谢。在本研究中,我们采用宏基因组分箱方法来探索参与纤维消化的单个微生物,并对瘤胃上皮细胞进行单细胞RNA测序,以研究促进VFA吸收和代谢的细胞亚型。
结果:本研究选取的52头泌乳中期奶牛(胎次=2.62±0.91)的产奶量为33.10±6.72 kg。利用宏基因组分箱法测定了186个细菌基因组的纤维消化和发酵能力,并鉴定出具有较强纤维素/木聚糖/果胶降解能力的特定细菌基因组,其与VFAs的生物合成高度相关。此外,我们基于20728个瘤胃上皮细胞的转录组,构建了包含18个瘤胃上皮细胞亚型的瘤胃上皮单细胞图谱。对编码VFA转运蛋白候选基因的表达谱的系统研究显示,IGFBP5+ cg样棘细胞独特地高表达SLC16A1和SLC4A9,表明这种细胞类型可能在VFA吸收中发挥重要作用。还确定了微生物组和宿主细胞之间的潜在相互作用及其在调节关键瘤胃上皮细胞亚型关键基因表达中的作用。
结论:本研究发现了在瘤胃中分别参与纤维消化、VFA摄取和代谢的关键微生物基因组和上皮细胞亚型。这些数据的整合使我们能够将微生物基因组和上皮单细胞与营养系统联系起来。
论文ID
原名:Investigation of fiber utilization in the rumen of dairy cows based on metagenome-assembled genomes and single-cell RNA sequencing
译名:基于宏基因组组装基因组和单细胞RNA测序的奶牛瘤胃纤维利用研究
期刊:Microbiome
IF:14.65
发表时间:2022.1.20
通讯作者:孙会增
通讯作者单位:浙江大学动物分子营养学教育部重点实验室
DOI号:10.1186/s40168-021-01211-w
实验设计

结果和讨论
1 宏基因组组装基因组(MAGs)的细菌分类组成
宏基因组测序从49头泌乳奶牛中共产生2,751,185,494条reads(补充表S1)。在去除低质量reads和宿主基因后,共产生2,697,595,628条clean reads,其中1,002,914,596条注释到RefSeq数据库,其中941,151,278条注释到细菌。将宏基因组测序数据(415 GB)进一步组装成34,039,290个contigs,用于重组MAGs。根据完整性和污染过滤MAGs后,从我们的数据集中总共获得186个瘤胃微生物MAGs,完整性>70%(88.22%±9.45%),污染<10%(5.81±2.71%)(补充表S2)。这些MAGs占宏基因组总reads的80.14%。所有186个MAGs的平均核苷酸同源性(ANI)均<95%,这意味着这186个物种是非冗余的。根据Parks等人提出的高质量MAGs定义,其中92个MAGs接近完整(完整性≥90%),94个MAGs基本完整(70%≤完整性≤90%);在这186个MAGs中,总共有71个MAGs显示出较低的污染度(污染<5%)。186个MAGs的基因组大小为659、879到26,548,065 bp(5,867,423±5,008,174 bp,平均值±SD),N50值为2108~61,696 bp。每个MAG的tRNA基因平均数为21.36±11.39%(范围从1~64),转移了11.84±4.43%的氨基酸(范围从1~20)。
然后对这些MAGs进行分类,186个MAGs被归为细菌(图1A),其中大多数MAGs属于未培养的谱系,没有任何先前的代谢或系统发育特征。79个MAGs可分为5个门:拟杆菌门(Bacteroidetes, 54个MAGs)、厚壁菌门(Firmicutes, 16个MAGs)、放线菌门(Actinobacteria, 7个MAGs)、变形菌门(Proteobacteria, 1个MAGs)和迷踪菌门(Elusimicrobia, 1个MAGs)。其余57.53%属于门的细菌MAGs缺乏现有的测序代表(图1A)。在目水平上,细菌MAGs被划分为6个目,包括拟杆菌目(Bacteroidales, 40个MAGs)、梭菌目(Clostridiales, 14个MAGs)、Coriobacteriales(5个MAGs)、双歧杆菌目(Bifidobacteriales, 2个MAGs)、Elusimicrobia(1个MAGs)和Selenomonadales(1个MAGs)(图1B和补充图S1)。在科水平上,将MAGs划分为6个细菌科:普雷沃氏菌科(Prevotellaceae,27个MAGs),红蝽菌科(Coriobacteriaceae,5个MAGs),双歧杆菌科(Bifidobacteriaceae,2个MAGs)、Elusimicrobiaceae(1个MAGs)、瘤胃球菌科(Ruminococcaceae, 1个MAGs)和韦荣球菌科(Veillonellaceae, 1个MAGs)(图1B和补充图S1)。我们对其中3个MAGs分类到种,包括Prevotella ruminicola、Selenomonas sputigena和Elusimicrobium minutum。
正如我们的研究和以往的研究表明,宏基因组分箱是一种无需培养程序即可恢复瘤胃微生物基因组(完全或接近完全)的有效技术。由于瘤胃的严格厌氧环境,基于培养的方法无法全面了解瘤胃微生物,该新工具使研究人员能够了解任何微生物组的功能,并易于有效地将瘤胃微生物与表型联系起来。为了充分了解瘤胃微生物在今后生产性状改良干预中的作用,将瘤胃微生物恢复到物种甚至菌株水平至关重要。然而,与之前的宏基因组分箱研究结果一致,我们的研究结果表明,大量未分类的微生物基因组和基因组无法解析到物种水平,许多MAGs仅在界水平恢复。以往关于瘤胃微生物群落系统发育多样性普查的研究也强调未分类细菌是瘤胃微生物中最丰富的。这些结果表明,仍有大量的微生物基因组有待测序和组装。

图1 基于宏基因组分箱分析,从49头奶牛中恢复了186个瘤胃MAGs。
A 186个瘤胃MAGs在门水平上的分类组成。B来自奶牛瘤胃的186个MAGs的系统发育树。彩色点:不同分类水平的MAGs;灰色点:分类到界水平的MAGs。MAGs:宏基因组组装基因组。
2 MAGs的CAZome谱
复杂植物纤维的分解是由瘤胃微生物的降解酶启动的,包括能够降解纤维素、木聚糖(半纤维素)和果胶的酶。我们基于宏基因组数据集和MAGs预测了CAZyme模块,以表征奶牛瘤胃中存在的碳水化合物活性酶。最常见的GH模块包括GH2(19,219)、GH13(15,765)、GH43(11,093)、GH3(6871)、GH97(5760)、GH31(5314)和GH5(5,308)(补充表S3);共鉴定出2964个参与纤维素降解的GH家族,包括GH5、GH9、GH44、GH45和GH48;共鉴定出13,110个果胶降解家族,包括GH28、PL1、PL9、PL10、PL11、CE8和CE12。参与木聚糖降解的CAZymes数量为24,488个,其中包括GH8、GH10、GH11、GH43、GH51、GH67、GH115、GH120、GH127、CE1和CE2。然后,我们将总宏基因组预测的CAZyme与公共数据库中的CAZyme进行了比较,图2A显示了总宏基因组预测的每个CAZyme模块的数量及其在CAZy数据库中的表示形式。
总体上,MAGs包含43968 GHs、3318 GTs、1612 CEs、328 CBMs、258 AAs和179 PLs(图2A)。图2B显示了MAGs中6个CAZyme模块的计数分布。平均而言,每个MAGs包含24.15±19.59 GHs(范围从0到112)、18.23±13.22 GTs(从0到72)、8.86±6.44 CEs(从0到30)、1.80±1.89 CBM(从0到8)、1.42±1.77 AAs(从0到11)和0.98±2.21 PLs(从0到15)。MAGs中蛋白质的分类和功能分布(图2C)显示未知细菌中CAZymes的数量最多,其次是拟杆菌门和厚壁菌门。CAZymes在细菌门中的分布(图2D)表明,GHs、GTs和CEs贡献了大多数门和未知细菌(不包括Elusimicrobia)中最多的CAZymes。CAZyme谱的表征突出了我们收集的不同微生物类群所使用的碳水化合物降解和利用策略。

图2 瘤胃MAGs的CAZyme谱。
A 在总宏基因组数据集和MAGs中预测的每个CAZyme模块的数量及其在CAZy数据库中的表示形式。B每个MAG编码的6个CAZyme模块的分布数量。C预测的CAZyme模块的门水平分类分布。D 6个CAZyme模块在每个门预测的CAZyme总数中的分布比例。MAGs:宏基因组组装基因组。
3 拟杆菌MAGs的纤维降解能力
我们的MAG数据揭示了拟杆菌门在瘤胃碳水化合物降解中的重要糖分解作用,因为这一优势门MAGs编码的蛋白质能够结合和消化多种碳水化合物底物。在我们的研究中,拟杆菌门MAGs编码了总GHs的38.85%(GHs是一组可以水解碳水化合物之间糖苷键的酶)。然后,我们分析了GH模块的分布,以及纤维素降解、果胶降解和木聚糖降解能力在54个拟杆菌MAGs中的分布(图3)。拟杆菌基因组中最常见的10个GH家族分别为GH2、GH5、GH13、GH97、GH25、GH28、GH78、GH105、GH106和GH23。此外,重要的纤维素降解CAZymes、果胶降解CAZymes和木聚糖降解CAZymes主要由属水平的普雷沃氏菌(Prevotella)(包括MAG234和MAG196)和种水平的P. ruminicola(包括MAG156、MAG361、MAG137、MAG278、MAG174、MAG231、MAG492和MAG214)编码(图3)。
CAZymes通常位于多糖利用位点(PULs)内,即编码细菌结合、转运和解聚特定聚糖结构所必需的酶的基因簇。拟杆菌门的分类群进化出PULs,GH模块最常在PULs中组织。由于PULs是特定于一个或多种底物,因此在MAGs中观察到的PULs可以用于预测微生物基因组的底物。然后我们在拟杆菌MAGs中检测到PULs,详细信息见补充表S4和补充图S2。从所有的拟杆菌MAGs中共鉴定出1827个PULs。在这些拟杆菌MAGs中,有50个含有一个以上的PULs,每个MAGs的PULs数量从1个到87个不等。PUL图谱显示,最常见的与CAZyme相关的PULs是参与木聚糖降解(GH43)、果胶降解(GH28)、淀粉降解(GH13和GH97),以及参与植物多糖消化和瘤胃微生物的碳水化合物降解(GH2、GH15和GH32)的辅助酶。
几个含有最丰富PULs的MAGs与P. ruminicola(MAG156、MAG361、MAG208、MAG27和MAG179,99% ANI)聚集在一起,这是一种厌氧革兰氏阴性菌,属于拟杆菌,可利用多种多糖作为底物(图3)。系统发育树的这一分支可能是含有新型多糖降解活性酶的微生物。在本研究(补充表S4)中,在P. ruminicola中检测到的PULs中存在的CAZymes主要是α-葡萄糖苷酶(EC:3.2.1.20)、阿拉伯内切-1,5-α-L-阿拉伯糖苷酶(EC:3.2.1.99)、β-呋喃果糖苷酶(EC:3.2.1.26)、β-半乳糖苷酶(EC:3.2.1.23)、内切葡聚糖酶(EC:3.2.1.4)、果聚糖β-果糖苷酶(EC:3.2.1.80),甘露聚糖内切-1,4-β-甘露糖苷酶(EC:3.2.1.78)、对硝基苄基酯酶(EC:3.1.1.-)和果胶酯酶(EC:3.1.1.11),表明这些MAGs可能具有较强的糖分解能力,并且可能能够适应多种饲粮条件下的瘤胃。Stewart等人报道,P. ruminicola是PULs的第二大贡献者(比P. multisaccharivorax含有的PULs较少),在该研究中,P. ruminicola PULs中编码的酶包括果胶裂解酶、果胶酯酶、葡萄糖苷酶、阿拉伯聚糖酶、半乳糖苷酶和甘露糖苷酶。本研究中最大的PULs来源(P. ruminicola)与Stewart等人(P. multisaccharivorax)的研究不同,可能是由于品种和饲粮的不同,因为品种和饲粮是影响瘤胃微生物群组成的两个关键因素。Stewart等人的MAGs是从三个杂交品种(包括Aberdeen Angus、Limousin和Charolais)和一个纯种(Luing)的奶牛身上恢复的,分别饲喂两种饲料,包括高精饲料(粗料:精饲料=75:925,基于干物质)和混合饲料(粗料:精饲料=480:520),而我们研究中的MAGs是从饲喂以玉米为主的高谷物饲料(粗料:精饲料=450:550)的Holstein奶牛产生的。在本研究中,瘤胃P. ruminicola拥有最多的PULs,这表明该分支具有很高的多糖降解潜力,这使得在Holstein奶牛的瘤胃中可以使用多种底物,并可能作为牛奶生物合成的重要中间代谢物贡献者。
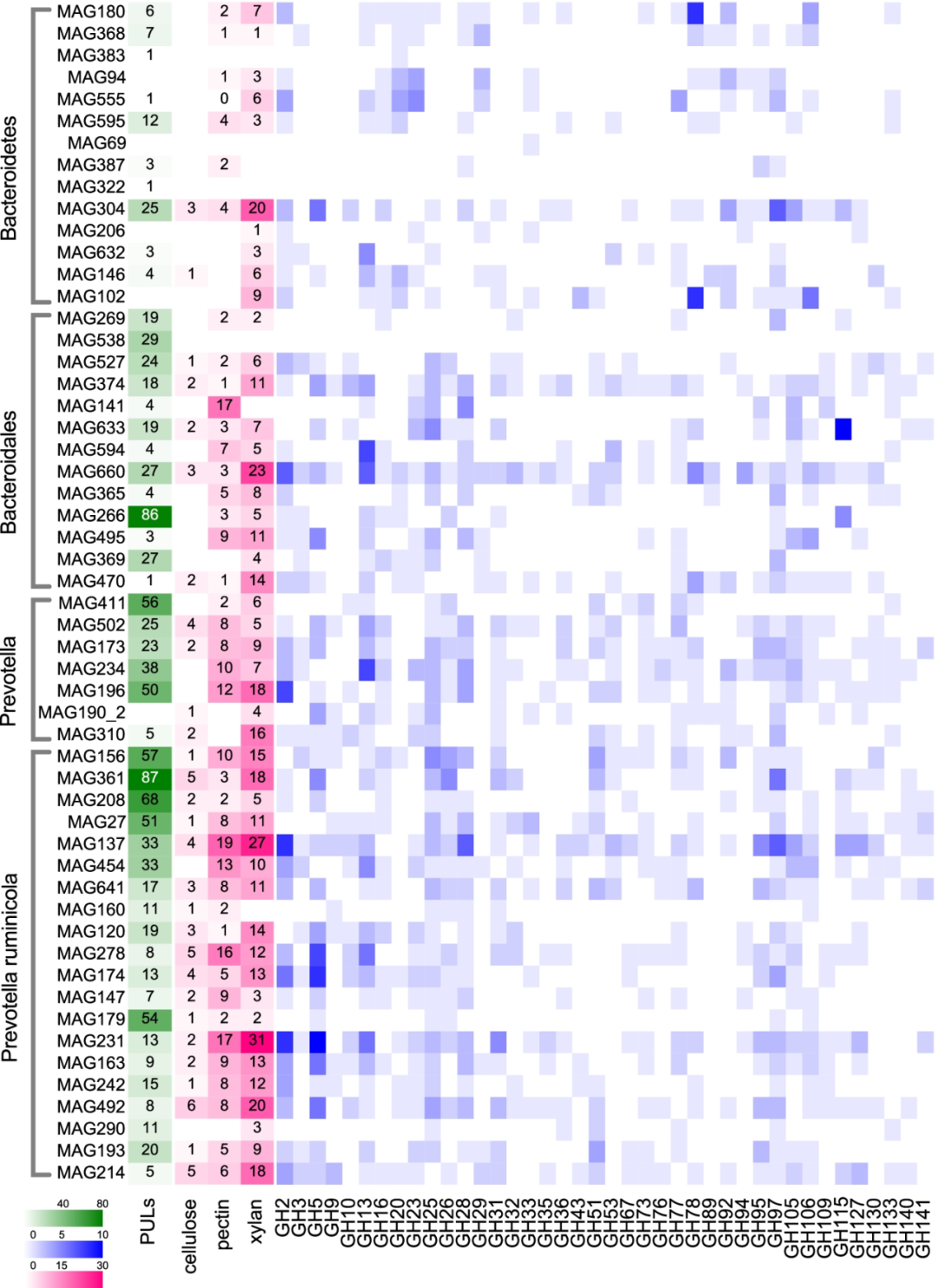
图3 拟杆菌MAGs的多糖降解潜力。
热图显示了MAG中编码的多糖降解CAZymes和GH模块数量,以及MAGs中多糖利用位点(PULs)数量。纤维素降解:GH5、GH9、GH44、GH45、GH48;果胶降解:GH28、PL1、PL9、PL10、PL11、CE8、CE12;木聚糖降解:GH8、GH10、GH11、GH43、GH51、GH67、GH115、GH120、GH127、CE1、CE2。
4 泌乳奶牛瘤胃上皮单细胞图谱
由于对瘤胃上皮细胞类型具有特异性的流式细胞术试剂的可用性有限,阻碍了体外功能测定的分选细胞,并且很难在体内直接研究瘤胃上皮细胞类型的功能,在瘤胃上皮中负责摄取VFAs的上皮细胞类型在很大程度上仍是未知的。scRNAseq的最新进展使单个细胞的表达谱得以实现,同时也是评估细胞类型特异性功能的间接手段。本研究中,我们对来自3头泌乳Holstein奶牛瘤胃腹侧组织的20,728个优质上皮细胞进行了scRNA-seq分析(图4A)。我们的结果显示,来自3头奶牛的细胞重叠良好(相似性统计分析R=-0.012和P=1)(图S4A-B),表明从这3头奶牛获得的数据保真度高,瘤胃上皮细胞图重现性好。我们总共在泌乳奶牛中发现了18个瘤胃上皮细胞簇(图4A)。细胞簇11、13、16为增殖的基底细胞(有丝分裂细胞,MC),表达标记基因KRT14、KRT5和MKI67,并根据其高表达的特异性基因,将其分为3个MC亚群(分别为TROAP+ MC、RRM2+ MC和MC_1)(图4B和图S5A)。细胞簇3、6、7和12高表达基底细胞(BC)标记基因KRT14或KRT5,并被定义为4个BC亚型(分别为KRT5+ BC_1、KRT5+ BC_2、KRT14+KRT5+ BC_1和KRT14+KRT5+ BC_2),特征是其高表达的特异性基因(图4B和图S5B)。
细胞簇0、4、5、8和14为颗粒细胞(GC)型,DLK2水平较高(图4B),其表达仅限于颗粒层,是角质形成细胞终末分化和角化的关键调节因子;通过比较,我们确定了这5个GC亚型中高表达的基因(图S5C)。细胞簇1、2和9被预测为三种棘细胞亚型(SC_1-3),因为它们不表达基底细胞标记物(如KRT14和KRT5),但轻度表达棘细胞标记物KRT10或颗粒细胞标记物DLK2(图4B),这表明它们可能是介于基底细胞和终末分化的颗粒细胞之间的棘细胞类型,我们也鉴定了它们的高表达基因(图S5D)。有趣的是,除了高水平的棘细胞标记基因KRT10、S100A8和KRT6A外,细胞簇10、15和17特异性表达GJA1(通道间隙细胞基因标记)(图4B和补充表S5);因此,将其定义为cg样SCs(通道间隙样棘细胞),免疫荧光染色进一步验证(图4C)。cg样SCs根据其各自的高表达基因进一步分为不同亚型,如TM4SF1+cg样SC(细胞簇10)、IGFBP5+cg样SC(细胞簇15)、BPIFA2C+ cg样SC(细胞簇17)与TM4SF1(P_adj=8.26e-90)、IGFBP5(P_adj=2.92e-251)和BPIFA2C(P_adj=4.04e-139)(图4B和补充表S5)。我们还发现这三种cg样SCs均高表达GSTA1和TMEM79(图S5E和补充表S5)。GSTA1编码一种保护细胞免受活性氧侵害的酶,而TMEM79在表皮完整性和屏障功能中发挥重要作用。脂肪酸结合蛋白基因FABP4(图S5E和补充表S5)在这三种cg样SCs中表达水平较高,说明它们参与脂肪酸的吸收、转运和代谢。

图4 泌乳Holstein奶牛瘤胃上皮单细胞图谱。
A瘤胃上皮单细胞的UMAP图显示瘤胃上皮细胞的18种细胞类型;细胞根据细胞类型着色。B瘤胃上皮单细胞图中各细胞类型的点图可视化。点大小表示一种细胞类型中细胞的百分比,颜色表示表达水平。C KRT6A与GJA1在瘤胃组织中的免疫荧光染色。比例尺:20 μm。每个组织独立重复两次实验,结果相似。UMAP:统一流形逼近和投影;BC:基底细胞;SC:棘细胞;GC:颗粒细胞。
5 负责VFA吸收的关键细胞亚型
阴离子交换机制在VFAs从腔内摄取到上皮细胞和排出到血液中发挥重要作用。依赖阴离子交换和不依赖阴离子交换的VFA转运有许多候选分子,包括SLC16A、SLC26A、SLC22A、SLC21A、SLC4A和SLC5A家族。我们分析了上述家族中基因在每种细胞类型中的表达,发现这些基因的表达模式具有细胞类型特异性,尽管其中一些基因(如SLC16A3、SLC16A7、SLC16A9、SLC16A11、SLC16A13、SLC26A3、SLC26A6、SLC22A7、SLC02A1、SLC5A8和SLC5A12)的表达百分比相对较低(图5A)。SLC16A9主要存在于BC亚型中,SLC26A2在MC_1、KRT14+KRT5+BC_2和IGFBP5+ cg样SC中检测到,但未发现显著差异。SLC4A7在GC_5中高表达(P_adj=4.46e-70)。KRT14+KRT5+ BC_2(P_adj=5.79e-150)、KRT5+ BC_2(P_adj=1.71e-248)、TM4SF1+ cg样SC(P_adj=9.67e-169)和IGFBP5+ cg样SC(P_adj=2.23e-281)高表达SLC16A1(图5A和补充表S5),它们在VFA吸收中起着至关重要的作用。此外,编码最有可能的VFA转运蛋白的SLC4A9在IGFBP5+ cg样SC中检测到高表达水平(P_adj=2.78e-77)(图5A)。SLC4A9也存在于KRT5+BC_2和TM4SF1+ cg样SC中,但未发现显著差异。这些结果表明VFAs可能很少被GC细胞亚型吸收,即使这些细胞是解剖上首先接触瘤胃腔产生的VFAs的活细胞。KRT5+BC_2、TM4SF1+cg样SC和IGFBP5+ cg样SC高表达VFA转运蛋白候选基因,并在VFA摄取中发挥更重要的作用。VFAs通过颗粒细胞到达基底细胞和棘细胞的机制有待进一步阐明。

图5 瘤胃上皮细胞各亚型VFA吸收和代谢的功能异质性。
A点图显示了所有瘤胃上皮细胞亚型中编码VFA吸收和代谢转运蛋白的基因的表达。节点大小表示细胞类型中细胞的百分比。从灰色到红色的颜色过渡表明基因表达水平由低到高。B利用VFA分解代谢过程和酮体生物合成过程基因集对瘤胃细胞亚型进行基因评分分析。VFAs:挥发性脂肪酸。
6 瘤胃上皮细胞亚型的VFA代谢异质性
VFA代谢是瘤胃上皮细胞利用VFA的另一重要功能。接下来,我们研究了瘤胃上皮细胞中与VFA代谢相关的基因(ACSS1、ACSS2、ACSS3、ACAT1、ACAT2、HMGCS2、HMGCL和BDH1)的模式。据报道,ACSS1、ACSS2和ACSS3可激活VFA代谢;但在KRT5+ BC_2和IGFBP5+ cg样SC中仅检测到ACSS2(图5A)。ACAT1在TROAP+ MC(P_adj=1.79e-177)、MC_1(P_adj=7.73e-126)、KRT14+KRT5+ BC_2(P_adj = 2.69e-108)和IGFBP5+ cg样SC(P_adj=1.80e-90)中以高百分比和高水平表达(图5A和补充表S5)。参与酮体合成的HMGCS2、HMGCL和BDH在KRT5+BC_2和三种cg样SC亚型中高表达(图5A和补充表S5)。综上所示,TROAP+ MC、MC_1、KRT14+KRT5+ BC_2、KRT5+ BC_2、TM4SF1+ cg样SC、BPIFA2C+ cg样SC,特别是IGFBP5+ cg样SC是VFA代谢的主要细胞亚型。基因集评分分析进一步证实了这一点,该分析表明,IGFBP5+ cg样SC在VFA分解代谢过程(μ=0.23)和酮体生物合成过程(μ=0.58)中得分均最高(图5B)。
7 MAGs和上皮细胞参与瘤胃纤维的利用
在瘤胃系统中,纤维的利用是通过微生物代谢和上皮细胞功能的相互连接来实现的。为了了解奶牛瘤胃微生物的碳水化合物降解活性和代谢潜力,以及瘤胃上皮细胞对VFA的吸收和代谢潜力,我们整合了我们的研究结果。图6A显示了关键细菌MAGs降解植物结构性碳水化合物和VFAs的潜力。基于与特定底物类别的联系,MAGs被分配到至少一个纤维解构和发酵途径。我们仅检测到用于解构难降解聚合物(纤维素、木聚糖和果胶)的强编码蛋白质的基因组包括MAG502和MAG174(纤维素);MAG304、MAG310和MAG660(木聚糖);以及MAG141和MAG454(果胶)。值得注意的是,几个基因组在编码一种以上的底物类型中发挥了重要作用。例如,MAG137(Prevotella sp.基因组)具有用于降解所有三种底物的强编码蛋白质,可以进一步研究从瘤胃中获取碳水化合物降解酶,用于动物饲料添加剂和基于木质纤维素的生物燃料生产。在利用中间代谢物(丙酮酸)产生VFAs方面,一些基因组表现出很强的能力,其中MAG403(参与乙酸和丁酸的生物合成)、MAG482和MAG73(参与丁酸和丙酸的生物合成)在多个VFA代谢途径中发挥作用。在此基础上,我们进一步了解了哪些微生物组编码瘤胃碳水化合物利用和代谢的重要功能。此外,该模型为我们理解这些微生物基因组中的基因功能,以及未来设计调控瘤胃微生物提高VFA产量和其他性状的策略提供了一个基本框架。由于这些基于宏基因组分箱的功能是潜在的,而不是真正的功能,因此未来需要进一步研究MAGs潜在功能背后的原因(例如,基于培养的调查)。
另一方面,VFAs的高效转运机制对动物生产效率至关重要。90%以上的VFAs以阴离子形式存在,被上皮细胞以不同方式从腔内吸收到血液中,尤其是通过蛋白介导的转运机制。与SLC16A1和SLC4A9不同,负责氯离子运输的SLC12A7在IGFBP5+ cg样SC中也有高表达(图5A和补充表S5, P_adj=3.00e-72)。以往的研究发现VFAs的摄取(尤其是丙酸)与氯离子的缺失呈负相关;因此,还提出了外向整流Cl-通道的参与。在IGFBP5+ cg样SC中,VFAs的细胞内分解(尤其是丁酸)可以维持VFAs通过瘤胃上皮的浓度梯度,以确保管腔摄取。综上所述,这些结果表明,这种上皮细胞类型可能在VFAs的摄取中发挥重要作用。

图6 提出的模型显示了细菌基因组的代谢潜力及其与瘤胃上皮细胞的相互作用。
A不同颜色的MAGs代表不同种类的微生物。左下方的示意图表示瘤胃上皮细胞的组成,右下方显示IGFBP5+ cg样SC中VFAs的摄取和代谢。B仅保留中间蛋白与靶基因之间具有转录调控相互作用的信号通路而获得的网络。
8 推测微生物群-宿主相互作用的功能效应
除了代谢物介导的微生物与宿主细胞之间的相互作用外,蛋白质-蛋白质相互作用是调节宿主细胞基因表达的最相关的分子相互作用类型之一。然而,在公开数据库中缺乏关于微生物组和瘤胃上皮细胞之间蛋白质-蛋白质相互作用的信息,因为检测种间蛋白质相互作用的实验技术既耗时又昂贵。在本研究中,我们使用MicrobioLink来识别微生物和牛蛋白之间潜在的相互作用,以及这些相互作用如何调节瘤胃上皮细胞亚型中关键基因的表达。首先,从上述30个关键MAG序列中获得223个微生物分泌蛋白(补充表S6)。利用UniProt对细胞膜和细胞外基质中的蛋白进行鉴定,共鉴定出8169个蛋白(补充表S7)。利用MicrobioLink中实现的结构域相互作用预测功能,共预测了涉及9个微生物蛋白和750个宿主受体蛋白的2949个相互作用(补充表S8)。由于宿主潜在的靶基因受到微生物-宿主相互作用的影响,我们重点研究了在IGFBP5+cg样SC和TM4SF1+cg样SC中显著高表达的前5个脂肪酸代谢相关基因(GJA1、GSTA1、FABP4、FABP5和ACSF2;avg_logFC=1.84和1.35;1.58和2.87;1.25和1.42;1.17和1.73;以及0.76和0.64;所有P_adj<0.05),以及与其他细胞亚型相比,在这两种细胞亚型(IGFBP5+ cg样SC中的CCDC80、AK1、CA12、PCK2和FPGS;以及TM4SF1+ cg样SC中的AGPAT2、ZNF750、CD82、BST1和ATP6V1B1)中高表达的前5个基因。随后,构建了介导受体向靶基因信号转导的潜在信号网络。为了保持特异性,排除了IGFBP5+ cg样SC或TM4SF1+ cg样SC中未检测到的宿主蛋白启动的信号通路。最后,我们获得了一个包含15个靶基因中的4个的网络,这些基因可能受到微生物组的调节(图6B)。在TM4SF1+cg样SC中,我们观察到一个调控BST1表达的特异性信号通路,包括CSNK1D、CDK5、STAT3、RB1和SPIL。该通路由MAG509中唯一发现的微生物蛋白标记基因1435调控。在TM4SF1+cg样SC中检测到编码CSNK1D基因的表达,但在IGFBP5+ cg样SC中未检测到(图6B)。MAG361、MAG528、MAG578、MAG509和MAG304参与调控信号网络中TM4SF1+ cg样SC和IGFBP5+ cg样SC中GJA1和FABP4的表达(图6B)。
在本研究中,微生物和瘤胃上皮细胞之间潜在的蛋白质-蛋白质相互作用是基于MAGs和RNA测序数据(上皮细胞)。未来的研究需要检测微生物和宿主细胞蛋白的表达,以进一步验证微生物和牛蛋白之间的相互作用,并解释这些相互作用如何调节瘤胃上皮细胞亚型中关键基因的表达。由于蛋白质-蛋白质相互作用是调节宿主细胞基因表达的最相关的分子相互作用类型之一,因此我们的综合分析有助于推断微生物蛋白通过信号网络对宿主细胞的间接影响,以及代谢物介导的微生物和瘤胃上皮细胞之间的相互作用。本研究中的所有数据都是公开的,为未来使用反刍动物模型的研究提供了一个比较框架。
结论
Holstein奶牛瘤胃微生物基因组的恢复为属或科水平上缺乏培养代表性的微生物基因组提供了新的见解。这一知识为建模和理解瘤胃微生物纤维分解提供了基础,以便从瘤胃中获取碳水化合物降解酶。此外,本研究的瘤胃上皮单细胞分析揭示了VFA吸收和代谢中的细胞类型异质性,其中IGFBP5+cg样SCs特异性参与这些功能。我们的数据提供了泌乳奶牛的基础知识,这使未来能够干预选择和控制微生物代谢和VFA吸收,从而改善反刍动物的生产性能和健康;但未来应考虑针对新发现的功能分析。
不感兴趣
看过了
取消

 打赏
打赏
不感兴趣
看过了
取消
精彩评论
相关阅读




中国医院排行榜















 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612



©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:82736610
 京公网安备 11010802020745号
京公网安备 11010802020745号


