
肠道微生物群免疫脑轴的异常发育可能在极早产新生儿的脑损伤中起作用。
编译:微科盟听雪斋,编辑:微科盟茗溪、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
早产儿患围产期白质损伤的风险很大。虽然肠道微生物群与早期生命发育有关,但对早产儿肠道微生物群免疫脑轴缺乏详细的了解。在此,我们分析了60名极早产婴儿的肠道微生物群、免疫学和神经生理学发育,这些婴儿接受了包括抗生素和益生菌在内的标准医院护理。我们发现严重脑损伤婴儿的皮质电活动成熟受到抑制。这伴随着γδT细胞水平升高和血管内皮生长因子T细胞分泌增加以及神经保护剂分泌减少。值得注意的是,肠道中的克雷伯菌Klebsiella过度生长可高度预测脑损伤,并与促炎症免疫张力相关。这些结果表明,肠道微生物群免疫脑轴的异常发育可能导致或加重极早产新生儿的脑损伤,并代表了新型干预策略的一个有希望的目标。
论文ID
原名:Aberrant gut-microbiota-immune-brain axis development in premature neonates with brain damage
译名:脑损伤早产儿肠道-微生物群-免疫-脑轴发育异常
期刊:Cell Host & Microbe
IF:21.023
发表时间:2021.9.3
通讯作者:David Berry & Lukas Wisgrill
通讯作者单位:维也纳大学;维也纳医科大学
DOI号:10.1016/j.chom.2021.08.004
实验设计
结果
维也纳大学医学院2017年9月至2019年6月间收治六十例极早产儿。研究队列的特征见表1和S1。在住院期间,监测神经生理发育,收集粪便样本,并在多个时间点抽取血液(图1A)。通过住院期间多个时间点的超声成像以及足月等效年龄时的磁共振成像(MRI)联合评估来确定脑损伤。在60名婴儿中,有7名死亡,38名显示年龄足够或仅有轻度脑损伤,15名婴儿被诊断为严重病理性脑损伤。观察到的损伤包括脑室周围出血性梗死(n=5)、脑室内出血(IVH分级>2,n=13)、小脑出血(n=7)和脑室周围白质软化(PVL,n=4)。这些病理中有许多伴随着脑容量的减少和蛛网膜下腔的扩大。IVH和PVL的代表性MRI图像如图1B所示。为了量化脑损伤,Kidokoro评分被用作脑损伤性质和程度以及受损脑生长的标准化MRI评估工具。正如预期的那样,严重脑损伤婴儿的Kidokoro评分显著较高(图1C;t检验;p=0.0001)。虽然胎龄(GA)被认为是脑损伤的一个重要预测因子,但我们发现出生时GA与神经影像学结果之间没有显著相关性(t检验;p=0.4371)。
为了描述患者的神经生理发育特征,我们采用振幅积分脑电图(aEEG)总分,该总分量化了四种不同的皮层电活动模式:(1)持续电压(CV),(2)不连续电压(DV),(3)突发抑制(BS)和(4)平坦跟踪(FT)。这些模式反映了测量时患者的皮质活动。我们观察到以初始静止期为特征的神经生理学成熟的一致趋势,其中DV是主要的aEEG背景模式(图1D、S1A和S1B)。随后是一段神经生理学成熟期(成熟期),在此期间CV持续增加,并最终在足月等效年龄(足月等效期)稳定下来。一般来说,脑损伤婴儿经历了延迟的成熟期,分娩后2周CV较低,BS较高(Wilcoxon;p分别为0.008和0.028)。此外,产后2周和4周脑损伤婴儿的DV显著降低(Wilcoxon;p分别为0.038和0.045)(图1D)。分娩后4周,神经生理成熟模式趋于一致,之后两组继续遵循类似的过程。
接下来,我们使用近红外光谱 (NIRS) 评估了脑组织氧合。为了量化脑组织氧合中潜在的有害不稳定性,我们计算了脑组织氧合 (cSO2) 极端波动的发生率,定义为cSO2在NIRS测量期间与平均值相差+/- 10 cSO2的时间分数 (VAR-cSO2)。我们观察到,在患有严重白质损伤的婴儿的早期成熟阶段(分娩后 4 周),cSO2的波动显著升高(t 检验;p= 0.025;图 1E)。
此外,我们评估了aEEG和NIRS信号在每个时间点对神经影像结果的预测潜力。我们发现,尽管平均cSO2对检测脑损伤没有预测价值,但VAR-cSO2在分娩后4周具有一定的预测能力(曲线下面积[AUC]=0.70)。BS 水平在分娩后 2 周最具预测性 (AUC = 0.80),通过结合BS和VAR-cSO2 (BS/100×VAR-cSO2),预测潜力在分娩后周 AUC = 0.79 并增加至 AUC = 0.74分娩后2 周和4周(图 1F和1G)。
因此,我们得出结论,神经影像学结果先于皮质电成熟的延迟开始。分娩后约2至4周,神经生理发育的短暂抑制和颅氧饱和度的不稳定性增加,可预测足月等效年龄的PWMI,并可能在其病理学中起重要作用。

尽管T细胞是肠道微生物组与大脑之间的重要联系,但对其早期个体发育及其与极早产儿围产期脑损伤的关系仍知之甚少。为了研究与围产期脑损伤相关的T细胞动力学的时间变化,我们通过荧光激活细胞分选(FACS)对40名极早产儿5个时间点的血液样本中的T细胞亚群及其相应的细胞因子反应进行了研究。其中,4名婴儿死亡,26名婴儿显示年龄足够或仅有轻度脑损伤,10名婴儿被诊断为严重病理性脑损伤。细胞因子反应的主坐标分析(PCoA)显示了3个不同的样本簇:一个早期生命簇(细胞簇1;分娩后平均天数:17)和两个晚期时间点的簇(细胞簇2和3;分娩后平均天数:分别为30和35)(图2A)。细胞簇2在患有脑损伤的婴儿样本中富集(细胞簇2为38%,细胞簇3为10%)。值得注意的是,与细胞簇3相比,细胞簇2在足月等效年龄时的Kidokoro评分以及结合VAR-cSO2的BS评分(BS/100×VAR-cSO2)更高(Wilcoxon;p分别为0.035和0.0088)。白细胞介素4、6和8(白细胞介素4、白细胞介素6和白细胞介素8)、血管内皮生长因子A(VEGF-A)和血小板衍生生长因子BB(PDGF-BB)的水平与观察到的PCoA差异显著相关,VEGF-A驱动细胞簇2的形成,其他细胞因子与细胞簇3相关。此外,我们还建立了分泌细胞因子和趋化因子的相关网络,以揭示其表达中的连接性。最值得注意的是,我们发现,与严重脑损伤的婴儿相比,从没有严重脑损伤的婴儿的样本中产生的网络显示出PDGF-BB更为核心的作用,后者的网络主要集中在白细胞介素 17a(IL-17a)(图S2A-S2C)。此外,通过使用细胞因子水平的地形数据分析,我们观察到早产儿出生后1个月的发育轨迹存在差异,这种差异在足月时趋于一致(图S2D)。
在比较有脑损伤和无脑损伤婴儿的细胞因子表达时,我们观察到有脑损伤婴儿的外周血T细胞在分娩后3天(Wilcoxon;p=0.048)和分娩后7天(Wilcoxon;p=0.034)的IL-17a和VEGF-A的生成增加,同时伴有白细胞介素22(IL-22)的降低分娩后7天(Wilcoxon;p=0.044;图2B)。尽管这些差异在分娩后28天消失,但分娩后28天脑损伤婴儿的样本显示神经保护剂脑源性神经营养因子(BDNF)(Wilcoxon;p=0.012)和PDGF-BB(Wilcoxon;p=0.026)的产生减少(图2B)。这些数据表明,出生后早期通过IL-17a的促炎性T细胞极化是后者异常多样化的基础,在此期间,VEGF-a反应失调是脑损伤发病机制的关键。图S2G中显示了量化的其他细胞因子和趋化因子,但在有脑损伤和无脑损伤的婴儿之间未显示出显著差异。
然后,我们通过FACS分析来自相同样本的调节性T细胞、辅助性T细胞、细胞毒性T细胞和γδT细胞(图2D-2J)。辅助性T细胞(TH:CD4+/CD3+)是最丰富的CD3+淋巴细胞,其次是细胞毒性T细胞(TH:CD8+/CD3+)、调节性T细胞(Treg:CD25+FoxP3+/CD3+CD4+)和γδT细胞(γδPAN/CD3+)(图2E)。脑损伤婴儿出生1周后T细胞水平升高(Wilcoxon;p=0.039;图2D),这与CD3+人群中γδT细胞水平升高相对应(Wilcoxon;p=0.027;图2F)。大多数γδT细胞以双阴性(DN;CD8和CD4)T细胞形式存在。出生后不久,约整个DN T细胞群中60%的细胞表达泛γδT细胞标记物。出生后1个月,DN PAN-γδT细胞的相对数量增加,且在脑损伤婴儿中明显增多(Wilcoxon;p=0.031;图2G)。此外,患有严重脑损伤的婴儿在32周纠正GA时的原始CD8+T细胞较少(Wilcoxon;p=0.031;图2H),在足月等效年龄时的效应CD4+T细胞较多(Wilcoxon;p=0.014;图2I)。尽管没有统计学意义,但我们观察到出生后不久脑损伤婴儿中迁移刺激CCR6+T调节细胞数量增加(图2J)。图 S2H 中显示了额外的 T细胞受体,这些受体被量化但在有脑损伤和没有脑损伤的婴儿之间没有显示出显著差异。
接下来,我们试图通过相关分析揭示VEGF-A、白细胞介素、神经保护剂PDGF-BB和BDNF与不同T细胞群的相互作用(图2K)。给出的值是从所有测量的细胞因子和T细胞的相关性分析中提取的。辅助性T细胞在所有时间点均与细胞毒性T细胞呈负相关,在分娩后3天和28天以及32周GA时与γδT细胞呈负相关。分娩后3天,VEGF-A与神经保护剂PDGF-BB和BDNF呈负相关。分娩后7天,辅助性T细胞与IL-10和IL-22呈负相关,而调节性T细胞与IL-1beta和IL-17a呈负相关。然而,γδT细胞与IL-8和PDGF-BB呈正相关。在分娩后28天,VEGF-A与γδT细胞的增加呈正相关,但与T调节细胞呈负相关。这种正相关性在以后的时间点消失,而与T调节细胞的负相关性持续存在。此外,我们通过典型相关分析(CCA)评估了足月年龄时细胞因子和趋化因子分泌水平,以及通过cMRI评估的Kidokoro评分和髓鞘形成评分对给定T细胞群方差的影响。我们发现,在所有样本中,调节性T细胞、辅助性T细胞、细胞毒性T细胞和γδT细胞的组成受IL-17a(p值:0.004)、IL-12P40(p值:0.004)、PDGF-BB(p值:0.002)和IL-4(p值:0.028)的表达以及足月龄时的脑髓鞘形成(p值:0.028)的显著影响,所有变量的综合解释力R2=0.26(图S2E)。
我们得出结论,分娩后 7 天开始 γδT细胞的增加可能与 PWMI 的发病机制有关。这种初始极化起初与 IL-8、IL-17a、VEGF-A 和 PDGF-BB 的分泌相关。随后,VEGF-A 和 γδ T细胞之间的密切联系持续存在,而神经保护剂 PDGF-BB 和 BDNF 的进一步分泌在患有严重脑损伤的婴儿中受到抑制。
图2.有无严重脑损伤的极早产儿的T细胞个体发育。(A) 主坐标分析(PCoA)-分离的细胞因子和趋化因子组成的双批次。轮廓评分确定了3个主要的细胞簇(由不同的符号表示)。符号的颜色(灰色到黑色)由分娩后的天数决定,婴儿年龄越大,符号越暗。符号的大小由Kidokoro评分确定,该评分通过cMRI在足月等效年龄评估,评分越高,符号越大。显著相关(p<0.05)的细胞因子/趋化因子绘制为箭头。此外,PCoA旁边的方框图显示了突发抑制和颅氧合变化(BS/100×VAR-cSO2)的组合值范围,以及各个细胞簇(细胞簇1,CC1;细胞簇2,CC2;细胞簇3,CC3)的Kidokoro评分范围。(B) 有(红色)和无(蓝色)严重BI的婴儿的血细胞因子/趋化因子浓度(产后3天,d3;产后7天,d7;产后28天,d28;孕32周,w32;足月等效年龄=足月)。(C) 说明FACS分析选通策略的代表性图像。不同颜色的门标记各个细胞群的选通。品红色,无目标;蓝色,辅助性T细胞;橙色,细胞毒性T细胞;绿色,T调节细胞;粉红色,γδT细胞。对于(D)和(F–J)中的方框图,深色表示患有严重脑缺血的婴儿的数据,浅色表示没有严重脑缺血的婴儿的数据。(D) 住院期间CD3+细胞的比例。(E) 调节性T细胞(绿色)、辅助性T细胞(蓝色)、γδT细胞(粉红色)和细胞毒性T细胞(橙色)的相对分布。(F–I)方框图显示在住院期间相对于给定父母群体的T细胞受体表达。(J) 方框图显示CCR6+/T调节细胞的表达,并添加由局部估计散点图平滑(黄土)产生的平滑线,以指示发展趋势。(K) Spearman相关分析,以揭示T细胞群与选定隔离的细胞因子/趋化因子在各自时间点之间的关联(p<0.001)。红色表示正相关,蓝色表示负相关。方框图显示组中位数和四分位数范围。星号表示p值:∗p<0.05,∗∗p<0.01,∗∗∗p<0.001。
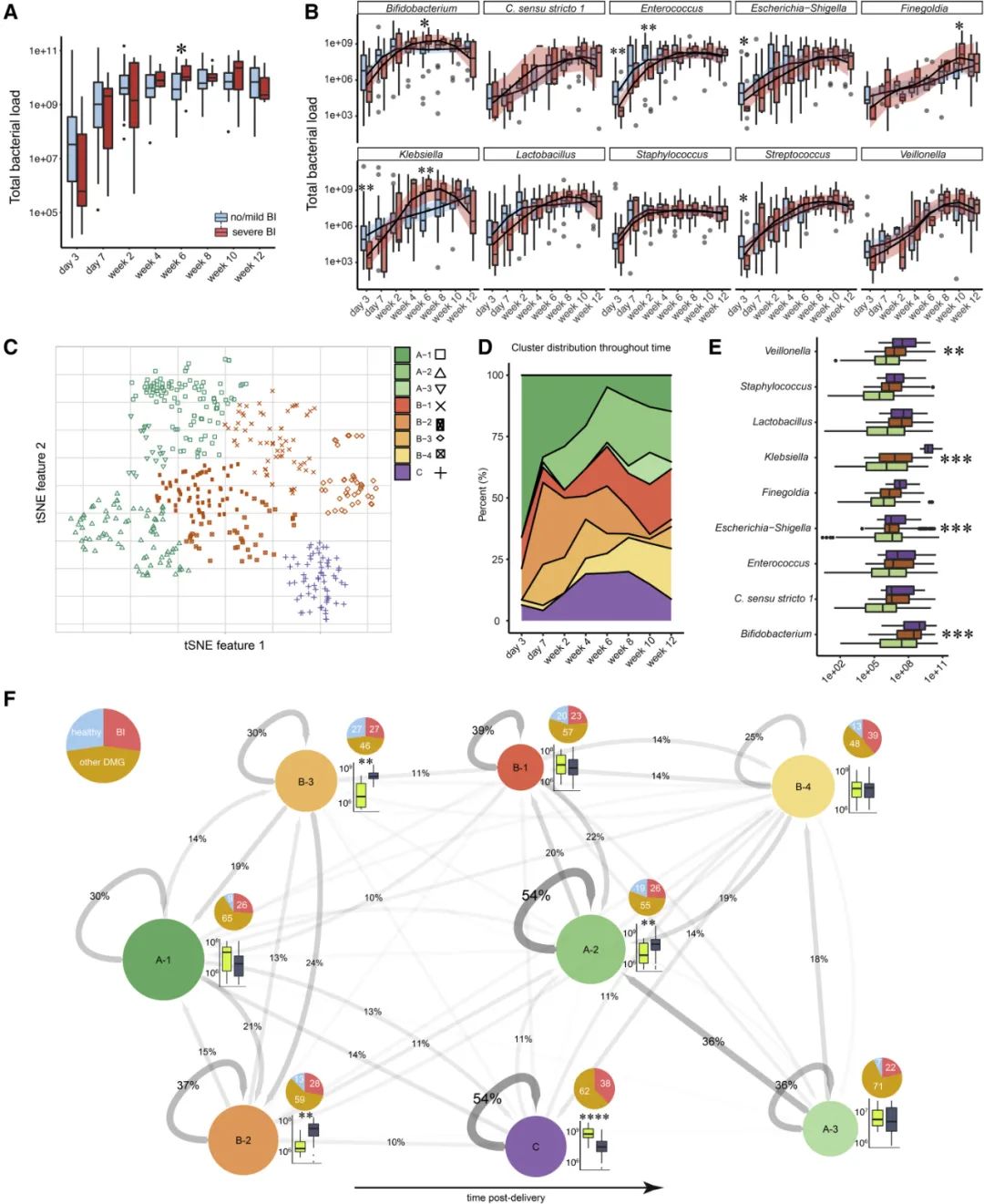
微生物群是免疫耐受的重要发起者,因为微生物抗原的性质和数量决定淋巴细胞反应。我们采用16S rRNA基因靶向的qPCR和16S rRNA基因扩增子测序来量化粪便样本中的总细菌负荷量,并对群落组成进行定量微生物组分析。我们观察到,随着时间的推移,细菌负载量增加,在分娩后2周趋于稳定,达到约每克粪便109个细胞(图3A)。分娩后6周,脑损伤患者的细菌载量略高(Wilcoxon;p=0.03),但除此之外,定植动力学与无脑损伤患者非常相似。总体而言,肠道微生物群主要由10个丰富且普遍存在的属(双歧杆菌、大肠埃氏菌-志贺氏菌、肠球菌、乳酸杆菌、葡萄球菌、链球菌、克雷伯菌、Clostridium sensu stricto 1、韦荣氏球菌和Finegoldia)组成,发生在至少20%的样本中,至少有105个细胞的丰度,并共同构成所有16S rRNA扩增子序列变体(ASV;图S3A)的75%。我们观察到严格厌氧核心菌 (Clostridia,Finegoldia和韦荣氏球菌)在产后早期很少出现,在后期大量增加(图3B;Pearson相关,R分别为0.53、0.62和0.48,p<0.001;图S3C)。令人惊讶的是,产后4周,克雷伯氏菌的数量平均增加了1.7倍(Wilcoxon;p=0.0049),但在严重脑损伤的婴儿中,产后不久,克雷伯氏菌的数量减少了(Wilcoxon;p=0.0066;图3B)。与这一观察结果一致,肺炎克雷伯菌ASV_c07_vnh_xuf与严重脑损伤之间存在显著的统计学关联(edgeR差异丰度检验;p=0.0467)。该菌株是从1名患有严重脑损伤的早产儿粪便中分离出来的,其基因组具有几种潜在的毒力因子以及使其能够在发炎的肠道内定植的基因,包括生物膜形成潜能、一系列抗生素抗性基因、铁清除铁载体以及异化硝酸盐还原为铵的基因(表S2和S3)。此外,我们观察到严重脑损伤的婴儿在分娩后6周双歧杆菌和出院前不久芬戈尔德菌的数量增加(Wilcoxon;p分别为0.0455和0.033)。然而,无严重脑损伤的婴儿在分娩后3天肠球菌、大肠埃氏菌-志贺氏菌和链球菌(Wilcoxon;分别为p=0.0083、0.028和0.045)水平升高,分娩后2周肠球菌数量增加(Wilcoxon;p=0.0065;图3B)。
为了更好地了解微生物组的组装和动力学,我们使用T分布随机邻域嵌入(tSNE)来识别不同的微生物组状态。该分析揭示了三个主要的微生物群,可进一步细分为八个子群,由轮廓评分聚类优化确定(图3C、3D和S3D)。总体而言,集群具有不同的细菌负载量(重复测量方差分析;p<0.001),集群C与集群A和B相比负载量增加(tukey事后检验;分别p=0.0017和0.0003)。集群A分为三个子集群,集群B分为四个子集群,而集群C没有形成子集群。此外,韦荣氏球菌属、克雷伯菌、大肠埃氏菌-志贺氏菌和双歧杆菌属在集群内的丰度存在差异(重复测量方差分析;p=0.002、<0.001、<0.001和0.001)。此外,与A组相比,C组中的维洛内拉菌升高(tukey事后检验;p=0.0012),与A组和B组相比,C组中的克雷伯菌升高(tukey事后检验;p=0.0012)≤ 0.001),A群中的大肠埃氏菌-志贺氏菌高于B群(tukey事后检验;p<0.001),C群中的双歧杆菌高于A群和B群(tukey事后检验;p分别为0.036和<0.001;图3E)。
接下来,我们使用这些子簇来检查微生物组动力学,并确定微生物组是否经历了可指示确定性演替的特征轨迹(图3F)。我们观察到,在生命的最初12周内,集群的频率有一种趋势,随着A2和B1患病率的增加,早期生命集群A1和B2逐渐变得不那么突出。在稍后的时间点,集群B4也变得更频繁地被检测到。簇A2和簇C具有最高的稳定性,即微生物组在随后的时间点停留在同一簇中的概率最高。在C群中,克雷伯菌是优势属,而在A2群中,大肠埃氏菌-志贺氏菌更为丰富。由于在C组中未观察到健康婴儿的粪便样本,我们得出结论,克雷伯菌在肠道中的优势和过度生长可能是驱动与神经病理学相关的独特且非常稳定的微生物组组装的关键。
此外,分娩方式和大量使用抗生素被认为是异常微生物演替的潜在驱动因素,尽管我们观察到患有和未患有严重脑损伤的婴儿在住院期间使用抗生素的时间没有差异,严重脑损伤与自然分娩有显著相关性(表1)。然而,抗生素给药的持续时间和给药方式与分类为C群微生物群的每位患者样本比例均无显著相关性(卡方检验,p=0.792和0.723)。这表明,自然分娩对病理性大脑发育具有特殊风险,但并不意味着随后会发展成簇状C样微生物群。
图3.极早产儿的微生物组群及其与脑损伤对应的多样性。(A)随着时间的推移,每克粪便的细菌细胞总数。(B)患有(红色)和未患有(蓝色)严重BI的婴儿中最丰富细菌属的绝对丰度。(C)通过绝对丰度和16S rRNA拷贝数校正的所有16S rRNA基因扩增文库的T分布随机邻居嵌入(tSNE)排序。层次聚类(Ward方法)确定了3个主要聚类(绿色、橙色和紫色)和8个子聚类(符号)。(D)显示子簇在整个时间内的百分比分布的面积图。(E)各主要集群中细菌属的绝对丰度。(F)网络说明了极早产儿微生物组演替的轨迹。圆圈对应于tSNE子簇。圆圈大小表示至少一次采样时在子群中发现的患者数量。箭头表示簇之间的过渡,箭头的厚度和阴影表示过渡的可能性。递归箭头表示同一簇内连续观测的百分比。饼图显示了被诊断为健康(蓝色,缺乏表1中列出的任何诊断)、严重BI(红色)或其他疾病(其他DMG,见表1;赭石色)的患者在各个集群中的分布情况。子簇根据簇中样本的平均患者年龄排列,从最年轻(左)到最年长(右)。圆圈旁边的方框图显示了相关子簇中克雷伯菌(亮绿色)和大肠杆菌志贺菌(深蓝色)的绝对丰度。方框图显示组中位数和四分位数范围。平滑线来自局部估计的散点图平滑(LOESS),并指示微生物群的发展趋势。星号表示p值:∗p<0.05,∗∗p<0.01,∗∗∗p<0.001。
虽然健康的成人肠道被认为是一个很大程度上缺氧的环境,但据报道早产儿的胃肠道部分有毒,因此在产后发育过程中必须过渡到缺氧状态。在缺乏末端电子受体(如氧)的情况下,细菌必须通过底物水平的磷酸化获得能量,而在肠道中,这通常是通过将膳食或宿主衍生化合物发酵成短链脂肪酸(SCFAs)来实现的,短链脂肪酸可作为神经系统和免疫系统信号分子。虽然肠道微生物群的主要营养来源很可能来自肠内营养,但我们观察到,细菌负荷与肠内营养量不耦合,在肠内营养量之前数周,细菌负荷趋于稳定(图3A和4A)。这可能是由于存在替代性基质,如胎粪、粘蛋白或产后肠细胞脱落。相反,胆汁酸、母体抗体和铁等替代性限制性营养素等分子可引入生长限制效应。
在特定取样时间点,我们未观察到患有和未患有严重脑损伤的婴儿在粪便中测量的醋酸盐、丁酸盐、丙酸盐、乳酸盐和甲酸盐含量方面的统计显著差异(图4B)。总的来说,SCFA的产量在分娩后约50天达到峰值,与细菌负荷不同,SCFA的产生似乎受肠内营养量的影响(图4C)醋酸和乳酸可以通过需氧代谢或厌氧代谢产生,但当细菌从需氧代谢转变为厌氧代谢时,细菌会将丙酮酸氧化为乙酰辅酶A和甲酸,而不是乙酰辅酶A、CO2和NADH。有趣的是,我们观察到分娩后1个月甲酸盐的累积量最高。随后丙酸盐和丁酸盐逐渐增加,这表明厌氧发酵活性随时间逐渐增加。专性厌氧类群的增加反映了这种向缺氧的转变(图3B和4B)。因此,我们收集了分娩前和分娩后28天的样本,以揭示氧影响和缺氧早产儿肠道的特征。分娩后28天,严格梭状芽胞杆菌1与丁酸盐的产生呈正相关,韦荣氏球菌属与乙酸盐和丙酸盐呈正相关(Pearson相关性;R分别为0.5、0.3和0.4,p<0.001;图4E),与其通过发酵生产乙酸盐和丙酸盐的能力一致。然而,由于我们在产后早期检测到少量丁酸和丙酸盐,我们得出结论,尽管出生后早期有氧代谢可能占主导地位,但发酵可能平行存在,可能存在于厌氧微生态位。为了测试早期或晚期发酵过程是否对脑损伤有影响,我们评估了SCFA与严重脑损伤的相关性。因此,我们计算了分娩前和分娩后28天样本库中每位患者的平均值(图4F和4G)。我们发现,在早期时间点,脑损伤患者的丁酸水平显著升高,丙酸水平略微升高(Wilcoxon;p分别为0.005和0.068),但在以后的时间点没有显著差异,表明早期向肠道缺氧过渡可能与脑损伤有关。
图4.极早产儿粪便中的SCFAs。(A)对于患有(红色)和未患有严重BI(蓝色)的婴儿,随时间进行肠内喂养。(B)患有(红色)和不患有(蓝色)严重BI的早产儿住院期间,nmol g-1粪便中的粪便SCFA。(C) 加权等高线图显示SCFA与肠内喂养百分比和产后天数的相关性。颜色越亮表示SCFA浓度越高。(D–F)(D)生后28天之前和(E)生后28天时或之后,细菌属丰度与SCFA水平[nmol]的Pearson相关性热图;生后28天之前(<d28)和之后(>d28)时,脑损伤(红色)婴儿和无脑损伤(蓝色)婴儿的SCFA浓度。(G) 在分娩后28天(<d28)和之后(>d28)有(红色)和没有严重BI(蓝色)的婴儿之间,总发酵量的差异以纳米/碳(nM/C)为单位测量。正方形表示组中值,误差条表示标准偏差。光滑的线条来自(LOESS),表明微生物群落的发展趋势。星号代表p值:*p<0.05,**p<0.01,***p<0.001。
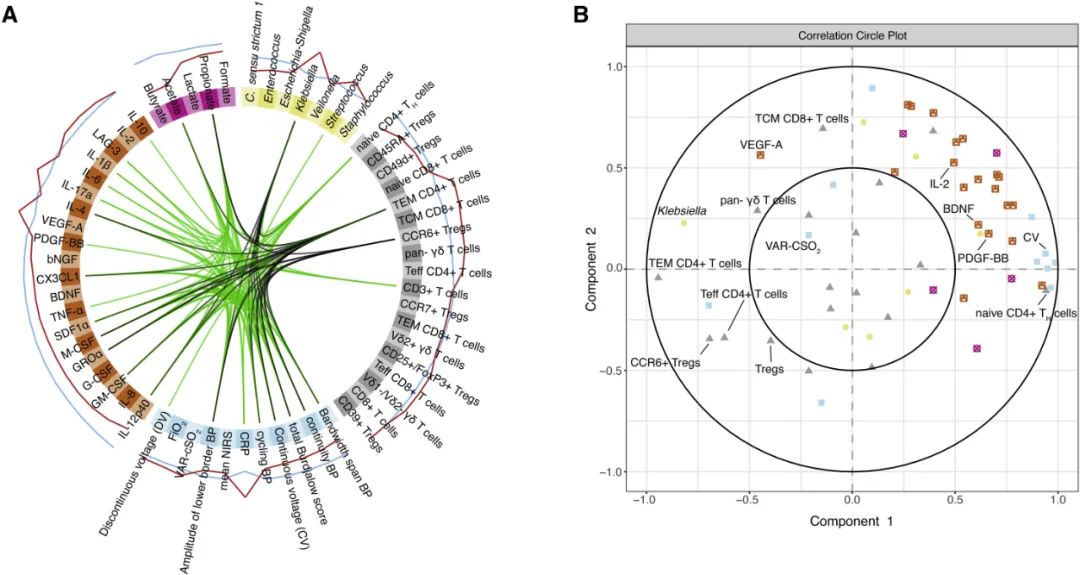
为了在我们的数据集中确定可靠的脑损伤生物标志物,我们对微生物组、代谢组、临床、免疫学和神经生理学数据进行了综合分析。白质损伤的最强指标是克雷伯菌水平升高、外周血C反应蛋白(CRP)、特异性T细胞群(效应记忆T细胞TH:Tem CD4+/CD4+、CCR6+T细胞受体和γδT细胞),以及几种细胞因子和趋化因子(IL-2、LAG-3、VEGF-A、IL-1β和IL-17a)的T细胞生成增加(图5A)。接下来,我们评估了不同数据类型之间的相关性(图5B)。我们观察到克雷伯氏菌水平与γδT细胞、VEGF-A分泌、CD4+T效应记忆细胞和CCR6+T调节细胞的相关性。同时,克雷伯氏菌水平与许多其他细胞因子/趋化因子的分泌呈负相关,包括神经保护剂PDGF-BB和BDNF。
总之,我们已经确定了一些提示肠道微生物群免疫脑轴发育异常的特征。患有严重脑损伤的婴儿表现出促炎性T细胞极化,其表现为外周血中γδT细胞的扩张,并在分娩后不久由升高的IL-17a和VEGF-a分泌驱动。随后,这些婴儿的神经生理学成熟开始延迟,其特征是皮层电位受到抑制,头颅氧饱和度增加不稳定性,以及神经保护剂PDGF-BB和BDNF分泌不足。肠道中的克雷伯菌过度生长是促使与严重脑损伤相关的独特且非常稳定的微生物组组装的关键。因此,我们得出结论,克雷伯菌参与肠道微生物群免疫脑轴的失调,并可能加重PWMI。
图5.极早产儿肠道微生物群免疫脑轴与脑损伤生物标志物。(A) 利用潜变量方法发现生物标志物的数据集成分析(Diablo)。整合微生物组(黄色)、代谢物(粉色)、T细胞(灰色)、细胞因子和趋化因子分泌(橙色)以及神经生理学/临床(蓝色)数据(n=84)。圆圈内的线表示绿色的正相关性,黑色的负相关性(典型相关性截止值:+/− 0.65)。圆圈外的线条表示年龄适当或轻度(蓝色)或重度(红色)BI的相应变量的预测潜力。(B) 相关圆图显示通过典型相关测量的变量之间的关联。相邻变量表示变量之间的相关性。
讨论
本研究涉及极早产儿肠道微生物群免疫脑轴的发育。我们观察到子宫外脑成熟的特征轨迹可分为三个主要阶段:神经发育静止期、神经生理成熟期和术语等效状态。利用系统生物学方法,我们发现肠道中的克雷伯菌过度生长可高度预测脑损伤(图5)。微生物群分析显示,以克雷伯菌为主的肠道菌群非常稳定,仅在患有疾病的婴儿中观察到(图3)。这种潜在的病理群体状态与静息期的促炎症和T细胞迁移以及神经生理成熟期伴随γδT细胞扩张的神经保护剂产生受损有关(图2)。
分娩后1周,我们观察到所有婴儿的CCR6+T调节细胞水平升高(图2J),表明免疫发育中出现迁移事件,部分原因可能是GIT中细菌数量增加(图3A)。脑损伤婴儿分娩后3天CCR6+T调节细胞水平较高(图2J),以及分娩后7天外周血中迁移刺激剂 VEGF-A 水平升高(图2B)。VEGF-A通过促进视网膜血管的异常分支在早产儿视网膜病变(ROP)的发病机制中发挥关键作用。有趣的是,所有脑损伤婴儿也被诊断为ROP(表1)。患有ROP但没有严重脑损伤的婴儿在外周血中显示出相似的 VEGF-A水平。然而,与患有严重脑损伤(Wilcoxon;p=0.0091)和患有ROP但无严重脑损伤(Wilcoxon;p=0.032)的婴儿相比,未受ROP或脑损伤影响的婴儿在分娩后7天VEGF-A水平显著降低。这表明分娩后早期VEGF-A分泌增加与ROP和脑损伤相关,尤其是严重的脑损伤包括ROP,但反之亦然。
此外,我们观察到严重脑损伤婴儿在分娩后7天和28天外周血中γδT细胞增加(图2F和2G)。γδT细胞是重要的早期免疫细胞,因为它们可以对先天性衍生的细胞因子作出反应,肠道内的γδT细胞已被证明能够感知微生物定植并提供免疫耐受和屏障功能。在促炎症状态下,γδT细胞部分从GIT组织中消失,并放弃其上皮内保护功能。IL-17a是γδT细胞向中枢神经系统迁移的重要信号,在中枢神经系统中,IL-17a会加重脑损伤。有趣的是,在我们的队列中,我们观察到出生后不久的患有脑损伤的婴儿外周血中IL-17a水平升高(图2E)。这些数据表明γδT细胞增殖和迁移至少部分由IL-17a驱动,并参与围产期脑损伤的发病机制。
肠道微生物群及其代谢产物具有显著的免疫调节潜力,包括例如能够驱动产生IL-17a的γδT细胞的增加。我们观察到,脑损伤婴儿在出生后的第1个月,粪便丁酸盐(一种主要微生物产生的代谢物)水平升高(图4F)。重要的是,粪便丁酸盐或其他SCFA水平的差异不仅由微生物生产量的改变引起,也可能由吸收和结肠细胞利用的改变引起。尽管丁酸盐对IL-10表达和上皮屏障功能有积极作用,但它也被证明会加重活动性炎症患者的GIT屏障功能障碍。由于早产儿可能损害上皮屏障功能,这些也可能是早期粪便丁酸盐水平升高的潜在原因,这可能会阻碍健康的粘膜发育。
据报道,在早产儿中,广谱抗生素处方和分娩方式会影响肠道微生物群的发育。鉴于自然分娩期间呼吸窘迫风险增加,可能导致脑损伤,剖腹产通常被认为是极早产儿更安全的分娩方法。这与我们观察到的自发性早产婴儿严重脑损伤发生率升高的情况一致(表1)。然而,剖腹产被证明能促进兼性厌氧菌的生长,如肠杆菌科,在我们的队列中,主要由大肠埃氏菌-志贺氏菌属和克雷伯菌属代表组成。有趣的是,我们观察到分娩后不久通过剖腹产分娩的婴儿中两种属的数量都有所增加(图3B),但分娩方式或抗生素治疗的持续时间与聚集性C样微生物群的建立之间没有关联,表明其他机制是特定病理社区状态发展的基础。肠杆菌科细菌在厌氧GIT条件下进行混合酸发酵,但在有氧条件下也通过有氧代谢旺盛生长。我们观察到,患有严重脑损伤的婴儿通常有较高的细菌负荷,分娩后6周的克雷伯菌数量也较高(图3A和3B)。与克雷伯氏菌相关的16S rRNA基因扩增子序列以及克雷伯氏菌分离物的基因组证据表明,肺炎克雷伯氏菌是该属的优势种(表S2)。最近已经证明,克雷伯氏菌能够利用早产儿肠道中的先锋菌群,并在微生物演替的后期建立,尽管其机制尚不清楚。克雷伯菌以前与败血症、坏死性小肠结肠炎(NEC)和新生儿医院感染有关。多重耐药和高毒力克雷伯菌已在全球出现,目前是严重感染的主要原因,尤其是在免疫功能低下患者中。强致病性克雷伯菌菌株可能具有致病因子,如粘附素、1型和3型菌毛、铁载体、厚荚膜和改变的脂多糖以逃避宿主识别。除了可能产生促炎细胞成分外,肺炎克雷伯菌的过度生长还可能直接通过黑暗发酵导致早期肠腔氧消耗以及导致早期生命丁酸水平升高或间接通过促进专性厌氧菌(如梭状芽孢杆菌)的定植(图3B)。通过这些途径,克雷伯菌可能驱动免疫细胞的促炎症极化,并负责随后γδT细胞的迁移。然而,目前尚不清楚克雷伯菌是否采用了特定的修饰,使宿主逃避达到过度生长,或者某些营养来源,如宿主来源的硝酸盐,是否在促炎症条件下可用,导致随后的水华,通过母乳衍生免疫球蛋白等机制压倒胃肠道清除。未来的研究应阐明克雷伯菌逃避肠道清除的机制,并通过鸟枪宏基因组学等方法揭示早产儿肠道中克雷伯菌菌株的多样性。
此外,益生菌作为一种潜在的抗肠道病原体的保护疗法被广泛讨论。事实上,益生菌处方似乎可以降低脓毒症和NEC的风险,但无法完全根除其发生。同样,我们发现队列中每天服用益生菌并不能完全预防PWMI。为了开发除广谱抗生素以外的先发制人措施,迫切需要更多集中于益生菌菌株在生命早期的宿主识别以及这些菌株与GIT中其他微生物群的相互作用的研究。
我们得出结论,肠道微生物群免疫脑轴的异常发育可能在极早产新生儿的脑损伤中起作用。具体而言,我们的数据表明,克雷伯菌过度生长以及相关微生物组的改变可能会加剧脑损伤,可能是通过触发免疫发育的变化,如γδT细胞水平升高。这些免疫改变,再加上随后神经保护剂的消耗,可能会影响神经生理发育和神经发育成熟。这些发现表明,针对肠道微生物群免疫脑轴的新治疗措施可能具有保护早产儿免于PWMI的潜力。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




