
调节性T细胞(Treg)因其免疫耐受作用,参与了肿瘤微环境的免疫抑制,是癌症靶向治疗的主要靶点之一。
调节性T细胞(Treg)因其免疫耐受作用,参与了肿瘤微环境的免疫抑制,是癌症靶向治疗的主要靶点之一。免疫代谢(Immunometabolism)在调节细胞状态和命运的作用机制越来越得到关注,Treg是否也受到免疫代谢所产生的异质性环境影响,而进一步影响其靶向治疗效果呢?2021年2月美国孟菲斯市圣犹大儿童研究医院免疫学系,迟洪波教授团队于《Nature》上发表了一篇《Lipid signalling enforces functional specialization of T(reg) cells in tumours》,阐述了脂质代谢重编程增强了肿瘤中Treg细胞的功能专一性。
1. 免疫代谢在调节细胞状态和命运中的基本作用已开始被阐明,但特定环境决定的代谢作用尚未得到充分诠释,特别是Treg细胞,其如何重新连接代谢程序以增强Treg细胞对肿瘤的功能适应。为了探索Treg细胞在肿瘤中的功能适应的分子机制,使用B16黑色素瘤接种WT小鼠后,对从肿瘤和瘤旁组织中分离出来的Treg细胞与外周淋巴结(PLNs)的作比较,进行了转录组分析。使用固化代谢途径的基因组富集分析(GSEA),提示:与PLNs相比,肿瘤中的Treg与脂质代谢相关的途径的富集度最高(Fig. 1a)。特别是固醇调节元件结合蛋白(SREBPs),作为促进脂类重新合成的转录因子,在GSEA中显示为高度富集的靶基因。遗传通路分析还显示,在瘤内Treg细胞的转录调节因子中,SREBP1(由Srebf1编码)和 SREBP2(由Srebf2编码)为上游顶部富集区。该富集区在小鼠黑色素瘤模型的公共单细胞RNA测序(scRNAseq)数据集中得到了进一步的证实,并在其他肿瘤细胞如乳腺癌、头颈部鳞状细胞癌中得到相似的结果。由此表明:在肿瘤微环境(TME)中的Treg细胞上调了SREBP基因。
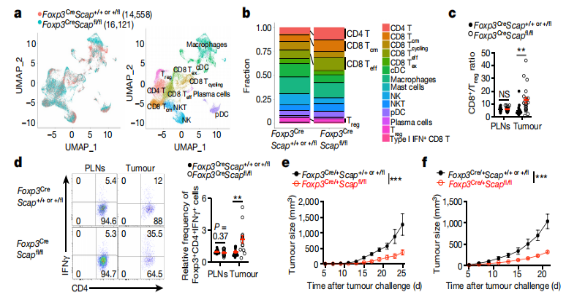
2. 为了确定SREBP信号通路在Treg细胞中的重要功能,使用Cre/loxP重组酶系统,建立了MC38腺癌小鼠及B16黑色素瘤小鼠模型的SCAP基因敲除鼠。SCAP(SREBP裂解激活蛋白)基因作为SREBP激活的必要启动子,被敲除后,明显抑制了小鼠黑色素瘤瘤内Treg细胞中SREBP基因靶点的表达。且Foxp3Cre Scapfl/fl小鼠中,B16黑色素瘤的生长显著减少(Fig. 1c)。为了探索SCAP/SREBP信号通路是否可作为肿瘤治疗的潜在标靶,研究人员亦建立了肿瘤小鼠中Treg细胞Scap基因的急性敲除模型,在成瘤后予以他莫昔芬和抗PD-1免疫治疗,在两种治疗方案下,SCAP敲除小鼠的肿瘤生长都有所减少,且对抗PD-1治疗尤为敏感(Fig. d-f)。以上结果提示:针对Treg细胞中的SCAP/SREBP信号通路的靶向治疗会释放出强大的抗肿瘤作用。
3. 为了确定SCAP缺陷Treg细胞在形成TME中的影响,研究者使用scRNAseq来描述B16肿瘤小鼠的CD45+免疫细胞(Fig. 2a)。与对照组相比,Foxp3CreScapfl/fl小鼠的TME中总/效应/记忆CD8+T细胞和总Foxp3-CD4+T细胞的比例增加(Fig 2),并通过流式细胞术分析进行验证。
4. CD8+T细胞与Treg细胞的比例是癌症相关的关键预后因子,在Foxp3CreScapfl/fl肿瘤中其比例也是增加的(Fig. 2c)。然而MC38结肠腺癌在Foxp3CreScapfl/fl小鼠中被排斥(Fig.1b),这样CD8+T细胞的耗竭导致部分肿瘤恢复生长,表明此效应部分依赖于CD8+T细胞的功能。
5. 此外,scRNAseq分析确定了Foxp3CreScapfl/fl小鼠肿瘤中Treg细胞的比例降低(Fig. 2b)。流式细胞术分析还显示,肿瘤中Treg细胞的占比降低,但其在Foxp3CreScapfl/fl小鼠的PLNs的占比没有降低,而瘤内Treg细胞的数量没有减少,可能是因为肿瘤浸润性淋巴细胞总数的增加。与肿瘤相比,在稳态下Treg细胞的积累没有变化。
6. 接着本文确定了SCAP缺乏对Treg细胞增殖状态的影响。与CD8+T细胞增殖中SCAP介导的胆固醇生物合成的要求一致,体外急性抗原刺激后,SCAP缺陷的Treg细胞显示有缺陷增殖,但这种缺陷可被胆固醇补充剂部分逆转。然而,Foxp3CreScapfl/fl小鼠肿瘤中的Treg细胞或PLNs并没有显示出胸苷类似物BrdU的变化,而SCAP缺乏对PLNs的Treg细胞也表现出由淋巴细胞减少诱导出增殖,这种现象同样也能被IL-2/抗IL-2复合物的治疗而引起。此外,与代谢驱动的增殖密切相关的CTLA4(细胞毒性T淋巴细胞相关蛋白4)的表达并没有因SCAP缺乏而改变。因此,SCAP功能在稳态和TME下对Treg细胞的增殖很大程度上是可有可无的,这突出了在CD8+T细胞中观察到的环境特异性增殖需求。
7. 检查Treg细胞的凋亡,发现SCAP缺陷的Treg细胞在肿瘤中活性Caspase-3(一种触发细胞凋亡的蛋白质)染色增加,但在PLNs中没有增加。然而,CD36的表达(调节瘤内Treg细胞生存所需的脂质摄取)和中性脂质的摄取不受SCAP缺乏的影响。同时检测了对照和SCAP缺乏的Treg细胞也显示出类似的线粒体轮廓,包括线粒体质量、膜电位和活性氧的产生。因此,SCAP对脂质摄取和线粒体适应性虽非必要,但有助于瘤内Treg细胞的生存。

8. 文献亦探讨了SCAP可能参与Treg细胞从静止的rTreg(CD44loCD62Lhi)细胞到激活的aTreg(CD44hiCD62Llo)种群的分化,以及Foxp3表达变化,这两者的表达都是炎症形成的条件。在稳态时,即使SCAP缺失,Foxp3的表达和肿瘤中aTreg的累积无论是数量和比例均无明显变化,所以SCAP在aTreg细胞累积和Foxp3表达变化上也非必要。
9. 在肿瘤内有一种特殊的“脆弱Treg细胞”,它表达Foxp3,但却存在分泌IFNγ障碍。研究发现,在稳态时,可分泌IFNγ的Treg细胞并未因为SCAP缺乏而发生改变;但与对照组相比,在SCAP缺失小鼠肿瘤内,能分泌干扰素的IFNγ+Treg表达增加(PLNs内却无此变化),且无论是B16黑色素瘤还是MC38结肠腺癌模型的肿瘤生长速度均受到显著抑制,这与脆弱Treg细胞在肿瘤中重塑TME的作用一致。
10. 对照组和Foxp3CreScapfl/fl小鼠的肿瘤浸润性Treg细胞进行转录组分析,SREBP在肿瘤Treg细胞中激活两种脂代谢通路, 包括脂肪酸合成和甲羟戊酸(mevalonate)代谢途径(Fig. 3a,b)。进一步的研究发现:Treg细胞中的FASN(脂肪酸合酶)缺失抑制了MC38和B16肿瘤的生长(Fig. 3c,d),但受SCAP缺失影响的仅TME。而FASN缺乏的Treg细胞中脂肪酸代谢途径下调。值得注意的是,由激活的TCRs(T细胞抗原受体)所驱动的Treg特异性基因也被下调了。假设FASN信号传递有助于Treg细胞的TCR依赖性激活和/或功能成熟。新分离的FASN缺乏Treg细胞在体外具有正常的抑制功能,由TCR诱导激活后,FASN缺陷激活的Treg细胞功能降低。为了将FASN与脂肪酸合成联系起来,体外添加了FASN的产物——棕榈酸盐,发现可恢复因FASN缺乏导致的Treg细胞激活障碍的作用(Fig. 3e)。此外,尽管有正常TCR诱导的增殖,FASN缺乏仍损害了TCR依赖的Treg细胞激活和成熟的标记上调。综上所述,FASN介导脂肪酸合成通路可促进瘤内Treg细胞的功能成熟,从而促进肿瘤生长。
11. SCAP/SREBP激活下游的其他机制也通过scRNAseq和流式细胞术进行分析,瘤内SCAP缺乏的Treg细胞降低其Pdcd1及PD-1表达显著降低,但在PLNs中没有降低(Fig. 3f)。相比之下,SCAP缺陷的Treg细胞在Treg特征分子CTLA4、ICOS、CD69、CD25、Nrp-1、CD39和CD73有正常表达。稳态下,Foxp3CreScapfl/fl小鼠的脾脏或NLT(非淋巴样组织)上的PD-1表达没有改变。因此,SCAP/SREBP信号是肿瘤中Treg细胞表达PD-1的必要信号,而在PLNs和NLT中则不需要。PD-1已被证明通过磷脂酰肌醇-3激酶(PI3K)的抑制信号,而PI3K的激活与Treg细胞的IFNγ产生的增加有关。抗PD-1治疗后,增加了Treg细胞中的IFNγ表达,且Treg细胞在肿瘤中pAKT和S6蛋白水平升高,但在PLNs中没有(Fig.3g, h)。而SCAP缺乏的Treg细胞在肿瘤中pAKT和S6水平亦升高,进一步支持SCAP和PD-1信号在瘤内Treg细胞PI3K活性中的作用(Fig. 3i, j)。SCAP/SREBP和PD-1抑制了肿瘤内Treg细胞中的IFNγ表达和PI3K信号传导。
12. 为了确定TME和其他背景对Treg细胞的PD-1表达的影响,并检查不同模型中PD-1的调节。与脾脏Treg细胞相比,TME Treg细胞诱导PD-1表达增加;然而,在EAE模型中并没有观察到Treg细胞诱导PD-1的表达增加(Fig. 4a)。在分子水平上SREBP信号是如何调控PD-1的表达,与上述表面蛋白的调节相一致,在肿瘤中,Pdcd1但不是Icos,其mRNA的表达降低,但在PLNs中没有发现差异(Fig. 4b),提示SREBP信号主要影响瘤内Treg细胞中Pdcd1表达。
13. 为了确定调节PD-1表达的上游信号,对从TME中分离出来的PD-1hi和PD-1loTreg细胞进行了转录组分析。PD-1hi细胞中Treg特异性的TCR依赖的基因及Nur77-GFP的表达升高(Fig.4d)。此外,体外TCR刺激后,PD-1在Treg细胞上的表达上调,同时也上调了SREBP基因靶点的表达,表明PD-1表达依赖TCR诱导。值得注意的是,Treg细胞中的SCAP缺失损害了TCR诱导的表面PD-1的表达(Fig. 4e)。因此,SCAP/SREBP信号传递将TCR刺激与Treg细胞上的PD-1表达联系起来。
14. SREBPs调节脂肪酸合成和甲戊酸代谢酶的表达。脂肪酸的合成对于PD-1的表达是不可或缺的,因为PD-1的水平在瘤内FASN缺乏的Treg细胞上不受干扰,甲戊酸代谢的参与,有助于合成胆固醇和异戊二烯类,且甲戊酸代谢有助于胆固醇和异戊二烯类药物的合成。通过代谢组学追踪示,SCAP缺乏的Treg细胞中醋酸衍生碳与胆固醇的结合有所减少(Fig. 4f)。针对甲戊酸代谢中的限速酶HMGCR的辛伐他汀,可以抑制TCR刺激的Treg细胞上PD-1的表达(Fig. 4g)。此外,HMGCR缺陷的Treg细胞在TCR刺激时显示PD-1表达有缺陷。然而,补充SCAP缺陷的Treg细胞的胆固醇,可部分纠正增殖缺陷。CTLA4的上调,不影响PD-1的表达。这些结果揭示了与胆固醇无关的甲戊酸盐代谢对Treg细胞PD-1调节的重要性。
15. 除胆固醇外,甲戊酸盐途径还分别生成类异戊二烯法尼基焦磷酸酯(FPP)和香叶基香叶基焦磷酸酯(GGPP), 这些脂质分子通过法尼基转移酶(Fntb编码的催化基因)和香叶酰香叶酰转移酶I型(Pggt1b编码的催化亚基)的酶活性调节法尼基化和香叶基香叶化的翻译后蛋白修饰。HMGCR下游的代谢物在辛伐他汀治疗的Treg细胞上的PD-1表达。(Fig. 4h)。值得注意的是,GGPP治疗也恢复了这些细胞上PD-1的表达,而FPP只显示出轻微的补救效果(Fig. 4h)。此外,甲戊酸盐或GGPP治疗也在很大程度上挽救了辛伐他汀治疗的Treg细胞中Pdcd1mRNA的表达。这些结果表明GGPP依赖蛋白香叶基香叶化在促进Treg细胞PD-1表达中的作用。
综上所述,SCAP的缺失会导致肿瘤组织而不是外周淋巴结中Treg细胞上PD-1表达减少,甲羟戊酸代谢介导的蛋白四异戊二烯化可增强Treg细胞中PD-1的表达。如此,SCAP/SREBP介导的脂质代谢通路对于Treg细胞特异性的在肿瘤微环境中发挥重要作用,为肿瘤免疫治疗提供了新的细胞和分子靶点,也再次强调了脂肪代谢在免疫系统中的地位。
编译:凌佳倩
审校:张军,缪长虹
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您





































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




