
全身麻醉(general anesthesia, GA)是一种药物诱导的可逆状态。

一、简介
全身麻醉(general anesthesia, GA)是一种药物诱导的可逆状态。包括意识消失、遗忘、镇痛、疼痛刺激反应消失和应激反应时血流动力学稳定这五个特点。然而全麻药机制一直是未解之谜。在麻醉的过程中,中枢神经系统中的神经活动已经在分子和脊髓水平得到了验证。GABA-A和NMDA受体似乎是最重要的靶点。全麻药阻碍了有害信息从脊髓上传至大脑。随着各种神经成像技术发展,大脑在不同意识状态下的神经活动和功能连接(FC)可用来研究全麻药机制。先前的研究表明,深度镇静剂量的全麻药优先减少高级信息处理区域活动,而对初级感觉皮层刺激反应没有影响;高级大脑网络中的皮质-皮质连通性降低,其中包括突显网络、默认网络(DMN)、执行控制网络(ECN)。虽在此方面研究有所进展,但业界还并未达成共识。
GA被认为是一种类似睡眠的行为状态,例如遗忘、镇静、可逆的意识消失。之前的研究认为,GA与睡眠状态都可能于意识消失的共同机制相关。例如,使用功能磁共振成像(fMRI)对静息状态网络的连通性研究表明,睡眠期间大脑皮层和丘脑皮质连通性模式的改变与全麻药诱导的GA相似。特别是在DMN和额顶网络(FPN)中发现了网内和网间的功能性分离。而DMN、FPN、初级感受器、丘脑皮质感受其之间的连通保持不变。但有其他研究提出,麻醉药诱导的意识消失与自然睡眠是由不同机制介导的。例如,研究发现丙泊酚诱导的意识消失与DMN网络中的PPC和楔前叶的连接保留有关;与丙泊酚和右美托嘧啶诱导的意识消失相比,自然睡眠状态保留了ECN与双侧顶上小叶的连通性。GA和自然睡眠共有大脑机制的假设还需要更多证实和详细的研究尽管如此,两者特征的比较可以帮助我们理解麻醉药对神经回路的影响和作用过程。
丙泊酚是最常用全麻药之一,其机制一直备受关注。有研究表明,PIS(丙泊酚诱导的镇静状态)与广泛的皮质网络中的区域性代谢降低有关,包括双侧额叶和顶叶联合区域的代谢降低,而感觉和运动皮质的代谢相对保存。在后扣带回(PCC)/楔前叶的研究也显示了异丙酚诱导的连接模式的改变。最近的研究表明,更高级别的静息状态网络(如外部控制网络和突显网络)参与了意识的改变。关于丙泊酚如何作用于功能网络的皮层连通性的研究取得了很大进展,但镇静机制及其与自然睡眠的异同仍不清楚。
图论网络分析已被广泛用于研究大脑功能结构。高度连接nodes是网络组织的一个普遍特征,这些nodes通常行使重要功能但也是弱点,一旦受到攻击可能导致整个系统快速崩溃。这些节点被称为rich-clubs,由高级别网络枢纽组成,在拓扑上高效率连接。rich-club通常在复杂系统的静态和动态过程中起着核心作用,对网络整体功能非常重要,因此受到了极大的关注。先前基于神经解剖学信息的网络结构研究强调了rich-clubs的重要作用,这些rich-clubs在暴露于不同的全身麻醉药(丙泊酚、七氟醚和氯胺酮)后会被破坏或是重组。在这些研究的基础上,我们假设(i)rich-clubs在丙泊酚诱导意识消失的过程进行重组(ii)rich-clubs改变可能主要分布在高阶认知网络中。为了验证这些假说,我们在药物诱导意识消失过程(包括清醒、丙泊酚诱导的轻度镇静【m-PIS】和深度镇静【d-PIS】)和生理过程(包括清醒和自然睡眠),构建基于fMRI的大规模脑功能网络。然后基于图论对PIS与自然睡眠的大脑网络的rich-club组织和节点属性(强度和效率)进行评估和分析。
二、研究过程
1、受试者
2015年12月1日至2016年11月30,招募了24名健康的右利手志愿者参与研究。12名容易入睡的志愿者分配至睡眠组,另外12名分配至PIS组:两组均为6例男性、6例女性。采用韦氏成人智力量表修订版(WAIS-RC)对所有受试者进行身心健康评估。所有受试者身体状况良好(BMI<30,ASAⅠ级)。以下情况不招募为志愿者:(i)过敏史、头部外伤手术史、药物成瘾史、哮喘史、晕动病史或在麻醉意外史;(ii)MRI检查的禁忌症,如血管夹或金属植入物、牙齿填充史或幽闭恐惧症;(iii)女性月经期或更年期。其中PIS组8例(男性4例,女性4例,平均年龄±SD:24.5±5.2岁),睡眠组9例(男性4例,女性4例,平均年龄±SD:23.5±3.8岁)。排除标准包括:(i)丙泊酚引起的烦躁反应导致剧烈体动;(ii)在预处理步骤中,头部最大运动3.0 mm和3.0°。表1列出了所有参与者的详细统计数据。此外,在数据采集之前,PIS组的参与者遵循标准的麻醉前禁食方案,并限制液体。实验结束后,受试者于完全康复后出院(麻醉出院后评分系统评分>9分)。
2、实验方案
所有参与者都在清醒状态下进行扫描,在放松状态下闭眼。PIS组的参与者使用TCI泵静脉注射1%丙泊酚。靶效应室浓度(Ce)设定为0.5~2.0μg/ml,间隔为0.5μg/ml。当达到预先设定的浓度时,暂停输药五分钟达到平衡,并使用fMRI进行总时长400s的扫描采集数据。同时采用Ramsay镇静评分(RSS)评估镇静状态。当RSS评分达到3分或4分时,认为受试者达到了m-PIS状态。当RSS评分达到5分或6分时,达到d-PIS状态。同时,在整个过程中连续记录所有受试者的生理参数,包括心率(HR)、血氧饱和度(SpO2)、平均动脉压(MAP)、呼末二氧化碳分压(PetCO2)。清醒、m-PIS、d-PIS之间HR、SpO2、PetCO2无显著差异,而d-PIS期间MAP显著低于清醒和m-PIS。因此,我们认为MAP参数在rich-clubs的统计分析中是一个没有意义的协变量。表1详细统计了生理参数信息。整个过程由两位有经验的麻醉医生负责给予丙泊酚和生理监测。
睡眠组的参与者没有给予任何药物,都在扫描仪下自然入睡。同时使用MRI兼容的脑电图(EEG)采集信号以评估睡眠阶段。一位脑电图专家为这项研究确定了睡眠阶段,提取并分析了非快速眼动(NREM)睡眠阶段2(N2)的fMRI数据。
3、数据采集
FMRI数据采集包括PIS组三种状态(觉醒、m-PIS和d-PIS)和睡眠组两种状态(觉醒和睡眠)。对于每个受试者,使用轴向三维快速扰相梯度回波序列(FSGPR)通过三维MRI序列获取高分辨率结构图像,参数如下:重复时间(TR)=8.876 ms,回波时间(TE)=3.516 ms,翻转角(FA)=13°,数据矩阵=256×256,视野(FOA)=256 mm×256 mm。功能成像采用EPI序列:TR=2000ms,TE=35ms,FA=90°,数据矩阵=64×64,分辨率=3.75×3.75 mm,层厚=5 mm,层间距1 mm。每个受试者共采集200容积,28个轴位切片,总扫描时间为400s,所有受试者都被固定,避免在无意识状态(自然睡眠和PIS状态)下头部或身体运动。
4、数据处理
使用统计参数图(SPM)和脑成像(静息状态)的数据处理和分析进行预处理。首先,时间序列中的前10个容积被排除,以避免MR信号中的非平衡效应。并且通过使用切片内插体素时间来校正功能图像的切片定时。接下来,对所有功能图像进行空间重新排列用于头部运动校正,并与其对应的解剖图像相互配准。然后将生成的图像与蒙特利尔模板进行空间标准化,并重新采样3x3x3mm3体素。使用半高宽为6 mm的高斯核空间平滑。因为先前的研究表明,网络波动在低频时最大。所以使用0.01-0.10 Hz带通时间滤波来最小化高频分量的生理噪声。尽管新出现的证据表明自发活动可能会在更高的频段(0.1赫兹以上)持续,背后的机制还没有定论。因此,我们的研究采用了典型的频率范围(0.01-0.10 Hz)。最后,回归出24个头部运动参数、全局信号(GS)、白质信号和脑脊液信号的潜在来源,以消除它们的影响。24个移动参数包括6个当前位置参数和6个过去位置参数,以及每个当前和过去参数的平方。
考虑到微运动对功能连通性的可能混淆效应,使用反映运动参数的时间导数的Jenkinson公式计算每个受试者头动情况(FD)值。排除平均FD>0.5 mm、平移>3 mm或旋转>3°的受试者。PIS组其余8名受试者(男4女)和睡眠组9名受试者(男5女4名)纳入进一步分析。
GS回归(GSR)是目前最具争议的静息fMRI预处理策略之一。GSR可以增加组织敏感度,减少运动依赖性,并增强整体神经元-血流动力学的一致性。然而,它也改变了组级的个体间差异。更重要的是,GS与神经活动有关且与所有受试者的脑电图警觉指标呈显著负相关。到目前为止,关于使用GSR的标准还没有达成共识。最近使用异丙酚的一项研究表明,与没有使用GSR的人相比,使用GSR后,在清醒状态下观察到更多的功能性大脑网络,对时间变异性进行了显著的任务调制。这些改善可能因为GSR可以进一步消除呼吸和运动等全局方差的非神经元来源。由于这些原因,我们的研究中也进行了GSR。
5、功能性脑网络的构建
Dos-160被认为是构建功能性脑网络的最佳模板,已在众多脑成像研究中得到应用,也有研究应用于脑中rich-club的研究。所以我们使用模板构建每个受试者的功能脑网络,选择160个感兴趣区域(ROI)(Fig. 1)该模板。160个ROI分成6个功能脑子网络,包括默认模式网络(DMN)、额顶网络(FPN)、带状盖网络(CON)、感觉运动网络(SMN)、枕部网络(ON)和小脑网络(CN)。首先对每个受试者通过体素的时间进程,提取160个ROI(球面半径为5 mm)的平均BOLD信号时间序列。然后,计算每个ROI的平均时间序列之间的Pearson线性相关性。然后,构造了160×160对称相关矩阵,并对对称相关矩阵进行了Fisher-z变换。此外,由于目前对负相关含义的含糊不清,所以在我们的研究中,计算绝对值矩阵,并将对角线的值设置为零。最后,根据先前对大脑网络进行基于图形的研究,用连接密度对连接矩阵进行阈值计算。
6、rich-clubs分析
6.1、rich-club组织
rich-club组织既可以在未加权的功能网络中进行分析,也可以在加权的功能网络中进行分析。在本研究中,我们采用加权网络分析丙泊酚诱导镇静和自然睡眠之间rich-club组织的变化。根据Dos-160地图集将加权网络节点定义为90个ROI,这些节点由带权重的边连接,表示两个区域之间的功能连通性。对于加权网络,在后续步骤中根据分组平均加权网络定义rich-club节点。首先,根据连通性权重对网络中的所有FCS进行排序,得到向量Wranked。每个节点到网络中其他节点的链路数计算为等级(K)。接下来,对于每个k值,我们选择等级大于k的节点集合,计算链接E>k的数量,并计算E>k连接的权重之和。加权rich-Club参数(k)被定义为W>K和E>K最强连接的权重之和之间的比值,后者来自于Wranked中最大连接数。由公式(1)计算得到ϕw(k)。
在每个加权网络基础上保持原有权重和等级生成l=1000个随机网络。每个随机网络中,对于每个k的等级,我们估算rich-club系数,由公式(2)得到。
先前研究报道,在连续的k范围内,(k)>1加权网络具有rich-club组织。
7、节点和连接的分类
根据组平均和稀疏网络选择rich-club成员组,来平均各组参与者的个体加权脑功能网络。与微观世界和无刻度组织的情况一样,rich-club组织的存在应该被视为整个网络的拓扑属性,当(k)>1时,rich-club组织与一定范围K等级的网络联系在一起。rich-club网络的边缘分类为三类连接:“rich-club”连接(连接rich-club成员)、“支线连接”(将rich-club节点链接到非rich-club节点)和“局部连接”(描述连接非rich-club节点的网络连接)。Fig2A提供了两类节点和三类连接的示意图。
8、节点属性分析
为了进一步探讨rich-club区域的重要性,计算和分析了节点特性,即节点强度和节点效率。它们的定义如下:
节点强度(NS)是节点所有连接边权重之和:
其中,Wij是节点i和节点j之间的边权重。它量化节点是否与图形相关。
其中,Lij是节点i和J之间的加权特征路径长度。
9、数据分析
本文中应用三种方法分析PIS组和睡眠组不同状态下的rich-club组织和结节特性(结节强度和效率)的差异:(i)PIS组镇静全过程(包括清醒状态、m-PIS和d-PIS)的单向重复测量方差分析(ANOVA)和作为正交分析的配对双尾t检验,以找出不同状态之间的差异,(2)用双尾配对t检验探查睡眠组中清醒和睡眠状态的差异,(3)采用双尾双抽样检验,以确定轻度/深度镇静与自然睡眠之间的显著变化。此外,包括年龄、性别和MAP在内的三个参数被认为是不感兴趣的协变量,并进行了回归以避免它们的影响。最后,将所有的p值设置为小于0.05,并设置错误发现率(FDR)校正来确定显著性水平。
三、结果
1、丙泊酚镇静组的rich-club分析
1.1、功能性脑网络中的rich-club
计算相关的rich-club参数,量化丙泊酚诱导意识消失过程网络rich-club组织。清醒状态、m-PIS状态和D-PIS状态下的群体平均功能网络显示了rich-club组织。不同状态下的rich-club系数曲线(Rnorm)如Fig2B所示。与清醒状态和m-PIS状态相比,d-PIS状态下的Rich-Club参数(k≤25)增大。与清醒状态相比,m-PIS状态下的Rich-Club参数(25≤,k,≤,30)略有降低。此外,与微观世界、无刻度组织的情况一样,rich-club组织的存在应该被视为网络的拓扑性质,并且rich-club组织与Rnorm超过1的k范围相关联。先前的研究显示,选择不同阈值的rich-club时能得到一致的结果。在本研究中,我们选择k=28的等级来研究丙泊酚引起的rich-club区域和在意识消失过程之间的联系改变。
2、丙泊酚镇静过程中rich-club分布
正如之前几项研究所描述的那样,rich-club区域是在组级别上选择的。三个不同状态K=28水平的rich-club数量和百分比如Fig2c所示。不同状态下,rich-club节点的百分比没有表现出任何显著的变化(清醒:16个,10%;m-PIS:17个,10.6%;d-PIS:19个,11.88%)。这说明在丙泊酚镇静的整个过程中,rich-club节点数是一致的。然而不同状态下大脑内部网络中rich-club的分布是不同的。如Fig2d所示,处于清醒状态的rich-club节点主要分布在DMN(50%)、FPN(12.50%)、CON(18.75%),ON和CN没有节点。m-PIS状态下节点主要分布在DMN(17.65%)、FPN(47.06%)、CON(5.88%)、ON(11.76%)和CN(17.65%),在SMN网络未观察到节点。在d-PIS状态DMN(26.32%)、FPN(36.84%)、CON(10.53%)、SMN(5.26%)、ON(15.79%)和CN(5.26%)。这些结果证明了丙泊酚诱导的意识消失过程中rich-club的重组。具体地说,部分rich-club区域在高级认知系统网络(DMN和FPN)和感觉系统(SMN和ON)之间切换。
为了进一步探索rich-club的重组,对rich-club区域的具体变化进行了研究。如Fig2e所示清醒状态下,这些节点主要分布在双侧角回、前岛、中岛、双侧扣带后皮质(PCC)、中央前回、楔前叶、背侧前额叶(dPFC)、额叶上叶、顶下叶(IPL)和颞回。然而在m-PIS和d-PIS状态下,rich-club节点主要分布在扣带回前部(ACC)、顶下叶(IPL)、顶下沟(IPS)、前额叶背侧皮质和小脑内侧。此外,部分rich-club节点(包括楔前、小脑内侧和枕叶)仅出现在m-PIS,而包括角回和枕后回在内的节点仅出现在D-PIS状态。以上结果显示,处于清醒状态的rich-club,包括额叶上叶、后扣带回皮质、前岛和中岛,首先受到丙泊酚的影响。当镇静致使意识消失时,紧随其后的是腹侧前额叶皮质、楔前叶以及部分枕前和中枕叶皮质的rich-club。此外,在丙泊酚诱导的意识消失过程中,包括小脑内侧区、顶叶下部和脑沟(IPL和IPS)在内的rich-club区域起着至关重要的作用。
3、rich-club、支线和局部连接的密度
根据rich-club和非rich-club的大脑区域分类,网络连接可分为三种拓扑类别:rich-club连接、支线连接和局部连接(Fig2A)。不同的状态下,三类连接的数量发生了显著变化(重复测量方差分析,rich-club,F=4.1160;p=0.0393;支线:F=4.1073,p=0.0395;局部:F=4.3986,p=0.0329),(详细见Fig3A)。与清醒状态相比,d-PIS(双尾配对t检验,rich-club:t=2.5118,p=0.0403;支线:t=2.4326,p=0.0452)中Rich-Club和支线连接数显著增加,局部连接数显著减少(T=−2.5061,p=0.0406)。d-PIS状态下的rich-club连接数显著多于m-PIS状态下的(T=2.3852,p=0.0485)。此外,局部连接强度也显示出相似的结果(F=10.2768,p=0.0018),(详细见Fig3B)。
与清醒状态相比,m-PIS状态(T=−2.3638,p=0.0500)和d-PIS状态(T=−4.5517,p=0.0026)的局部连接强度明显降低。此外,与m-PIS状态相比,d-PIS状态下rich-club连接的强度显著增加(T=−2.3942,p=0.0479)。这些结果提示,丙泊酚诱导的意识消失过程中,局部连接有向rich-club连接转变的趋势,内部rich-club组织以及rich-club与非rich-club组织之间的沟通加强了,提示在丙泊酚诱导的无意识状态下,rich-club组织与非rich-club组织之间的连接有向rich-club连接转变的趋势。
此外,这三种分类的连接在功能性大脑网络中的分布也发生了改变(Fig3C)。在清醒状态下,网内rich-club连接主要位于DMN,少数位于FPN、CON和SMN。网间rich-club连接主要分布在DMN、FPN、CON和SMN之间。然而m-PIS状态下,网内rich-club连接转移到了FPN和CN,而网间rich-club连接位于的FPN、CON和CN。最后,d-PIS状态下的rich-club连接是DMN和FPN之间的模块内连接和模块间连接。同时,我们发现,对于12%、15%和18%的成本,都具有较好的稳健性。这些结果表明,丙泊酚确实影响了rich-club连接的空间分布。在丙泊酚诱导的意识消失过程中,FPN的网络内连接和FPN与其他功能网络之间的网络间连接更多地出现在rich-club连接中。
4、自然睡眠人群rich-club分析
4.1、自然睡眠期间rich-club分布
睡眠组采用与PIS组相同的rich-club分析过程。重要的是,为了在PIS和睡眠组之间进行公正的比较,睡眠组中rich-club组织设定k=26等级,以获得相似数量的rich-club区域(Fig4A)。对于睡眠组,在清醒和睡眠状态下,rich-club节点的数量和百分比是相同的:分别为17%和10.63%(Fig4B)。然而,rich-club节点在两个状态的分布是不同的。特别是在清醒状态下,节点主要分布在DMN内,其余均分布于FPN(11.76%)、SMN(5.88%)、ON(11.76%)和CN(11.76%)。睡眠状态下,节点主要分布在DMN(29.41%)、SMN(35.29%)和ON(17.65%),其他主要分布在FPN(11.76%)和CON(5.88%),(详细见Fig4C)。此外,清醒时的rich-club节点位于扣带回的前后部、角回、IPL、IPS、小脑外侧区、PCC、楔前叶和枕回;睡眠状态的rich-club节点位于角回、额叶背侧和上额叶、中岛、PCC以及部分枕叶和颞叶(Fig4D)。这些结果显示,在自然睡眠诱导的意识消失过程中,rich-club区域从高阶认知系统网络(DMN)和小脑系统(CN)转移到感觉系统(ON和SMN)。结合PIS组中rich-club组织的结果,我们的研究结果表明,在丙泊酚和自然睡眠引起的意识消失过程中rich-club确实发生了重组。然而,rich-club区域变化模式略有不同。
5、rich-club、支线和局部连接的密度
清醒和睡眠状态之间不同类别连接的数量和强度没有显著差异(p>0.05)(Fig5A、B)。然而,与清醒状态相比,睡眠状态下rich-club连接的分布发生了明显的变化。在清醒状态下,DMN的网内连接以及DMN、FPN和CN之间的网间连接存在rich-club连接和支线连接。在睡眠状态下,rich-club连接位于DMN、FPN、SMN、CON和ON的内部和互连网络中。这些结果表明,睡眠状态下不同类别的连接没有整体变化。然而,三类连接的空间分布是不同的。
6、丙泊酚诱导镇静与自然睡眠的连接改变
为了进一步研究不同类型意识消失过程的变化,比较了镇静状态(m-PiS和d-PiS)和睡眠状态(Fig6A、B)中不同连接类别的数量和强度。与睡眠状态相比,d-PIS状态下rich-club连接的数量和强度显著增加(双尾两样本检验,t=3.3887,p=0.0041;强度t=3.0426,p=0.0082)。此外,局部连接的数量和强度明显减少(数量:t=−2.5037,p=0.0243;强度:t=−4.4614,p=0.0005)。除局部连接强度外,m-PIS和睡眠状态之间无显著差异(T=−2.9496,p=0.0099)。结合上述结果表明,功能性脑网络中rich-club重组模式可以区分异丙酚诱导和睡眠诱导的意识消失。
7、关于局部拓扑性质的分析
7.1、PIS状态下节点强度和效率降低
我们进一步研究了异丙酚引起的rich-club区域和非rich-club区域在意识消失过程中局部变化。在图论和网络分析中,加权网络的基本和常见拓扑性质是节点强度和节点效率。之前的研究表明,rich-clubs在整个大脑网络中发挥着核心作用,并对该网络效率有很强的积极影响。因此,我们确定了在局部拓扑属性上表现出显著组间差异的脑区,包括节点强度和节点效率。与清醒状态相比,在m-PIS和d-PIS期间,右侧中岛、双侧颞回和左侧中央前回的结节强度明显降低,而左侧前额叶皮质的结节强度显著增加。与m-PIS状态相比,d-PIS状态下左侧中央前回的结节强度明显高于m-PIS状态,(详细见Fig7A)。与清醒状态相比,PIS状态下的左侧腹内内侧PFC(VmPFC)、左侧前岛(AInsula)、右侧中岛、右侧额回、左侧中央前回、右侧顶回和双侧颞叶的节点效率显著高于清醒状态。此外,与清醒状态和m-PIS状态相比,d-PIS状态下左前额叶腹侧皮质(VFC)的结节效率明显降低(Fig7B)。经FDR校正后,显著改变区域的节点拓扑属性的p值均小于0.05。
8、自然睡眠状态下节点强度和效率的变化
我们还研究了睡眠状态下功能性脑网络局部拓扑性质的变化。与清醒状态相比,睡眠状态下右侧辅助运动前区(PreSMA)和右侧楔前脑区的节点强度明显降低,而左侧顶叶和颞回的节点强度则显著增加。右侧preSMA和楔前叶的节点效率明显降低,而左侧颞回的节点效率明显升高(Fig8A、B)。
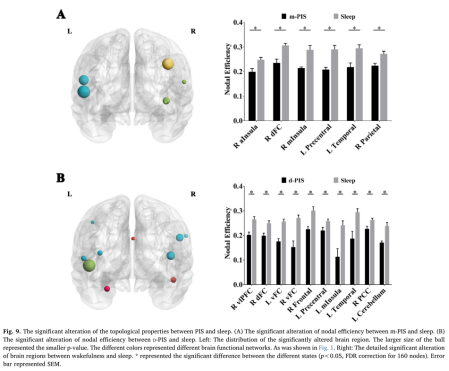
9、与自然睡眠相比,PIS状态下的节点效率降低
与睡眠状态相比,在m-PIS状态下,右侧前中岛、右侧额叶背侧、左侧中央前回、左侧颞回和右侧顶回的节点效率显著降低。右侧腹侧前额叶皮质(VLPFC)、右侧DFC、双侧VFC、左侧中央前叶、左侧中岛(MInsula)、右侧PCC、左侧颞叶和左侧小脑的节点效率也显著降低(Fig9A、B)。
四、讨论
rich-club组织表现出跨越许多不同大脑区域的复杂网络关系,并在整个大脑中执行最有效的信息传输。了解丙泊酚诱导以及自然睡眠的意识消失过程中脑功能网络rich-club结构的变化,将为理解丙泊酚的神经生物学机制带来新的见解。在本研究中,利用rs-fMRI构建了功能性脑网络中的rich-club组织。这些结构随后在两种相似行为反应状态之间进行了比较:在异丙酚由浅入深镇静和自然睡眠。这一些有趣的发现可以解释先前GA机制研究中报告的临床和实验结果。
4.1、丙泊酚诱导镇静过程中rich-club重组
rich-club在整个网络结构中扮演着中心角色。攻击rich-club节点对大脑网络功能伤害是其他随机位置的三倍。这些结果表明,大脑中的整体传递主要由rich-club组织控制。同时,rich-club区域在远距离脑通讯中的核心作用与功能连通性研究结果一致,功能中枢在全脑通讯效率以及认知功能方面发挥着重要作用。在脑功能领域,我们的研究首次提出,丙泊酚逐渐引起的意识改变对rich-club组织有不同程度的改变。值得注意的是,在逐渐消失行为反应性的过程中,rich-club的联系并没有中断,而是重组。结果表明丙泊酚诱导意识消失的机制与脑中枢在功能网络中的重新分布密切相关。
4.2、丙泊酚诱导镇静过程中rich-club的变化规律
我们的研究表明,丙泊酚诱导意识消失过程中rich-club区域在高级认知网络(DMN和FPN),感觉和运动网络(SMN和ON)和小脑网络(CN)之间转移。连接性减弱和丙泊酚引起的意识逐渐减弱之间的相关性在大多数默认网络和执行控制网络的领域普遍存在,这与之前的研究结果一致。相反,在早期的视觉和听觉皮层中,没有发现连接性和意识之间的关系。此外在丙泊酚的影响下,大量的局部连接转换到rich-club连接。我分别从rs-fMRI和EEG证明了意识改变时DMN和FPN脑区的拓扑参数被破坏。还有一项研究也证实了先前的发现,暗示了FPN和DMN的参与,强调了网络级拓扑中可能在调节意识状态的过程中产生了至关重要的作用。研究者认为FPN在认知控制和适应性执行任务发挥了核心作用,还揭示了FPN在中央执行网络和默认模式网络之间转换的关键作用。此外,FPN可能控制负责控制复杂运动的协调和规划。基于EEG的GA研究显示,PIS期间额顶连接中断。丙泊酚的不同程度镇静降低听觉皮层对声音刺激的反应,甚至大量丙泊酚诱导使警觉性反应消失时听觉反应仍然存在,但对于不同声音进而词汇刺激地特异性反应消失。警觉程度取决于大脑皮层之间信息地有效变化、交换和整合。我们认为中枢从高级认知网络和感觉网络到运动网络地转变和重新分布可能使丙泊酚诱导意识消失地重要特征。
在PIS期间,小脑网络中rich-club区域地数量也发生了显著变化。传统认为小脑与运动功能有关,接受来自脊髓或皮质和皮质下区域感觉系统的输入,并整合这些输入信息来调节运动活动。小脑参与各种功能,包括运动语言控制、眼球运动功能、握力、随意肢体运动和经典条件反射。小脑还参与听觉意识、详细记忆提取和特质焦虑。动物研究也证明了小脑在维持意识方面的重要性。小脑-大脑连接调节可能是意识水平改善的迹象。研究表明,在异丙酚诱导的镇静和恢复期早期,小脑活动减少。从本质上说,小脑可能在维持清醒方面起着重要作用,因此可以成为全身麻醉药的主要靶点。
3、rich-club组织在丙泊酚诱导镇静的过程中加强了连通
一项研究发现,少量的rich-club区域之间的连通性却主导了整个大脑网络的聚集,揭示了rich-club连通性在整个网络中发挥着杠杆作用。在一个非常小rich-club中调节连接模式就足以产生一个具有同配型或传递性的网络。这些结果表明,rich-club连接在脑网络交流中起着重要作用。在我们的研究中,rich-club连接的数量和强度受到丙泊酚的调节。尤其是在d-PIS过程中,与清醒状态相比,rich-club连接的数量和强度显著增加,而局部连接的数量和强度显著降低。此外,在m-PIS和清醒状态下,rich-club、支线和局部连接的数量和强度没有显著差异。我们的研究结果表明,在丙泊酚诱导的意识消失过程中,需要更多rich-club连接来促进整个大脑的交流。此外,“内在连通性网络”的定义有助于理解丙泊酚麻醉的可能作用靶点。这些网络由DMN、执行控制网络(ECN)、视觉和听觉网络组成。是更高级的信息处理网络,产生心理活动和自我意识。我们发现,在丙泊酚诱导的意识消失,rich-club连接性的分布正在逐渐转变。特别是,高阶认知网络(DMN和FPN)的网内rich-club连接显著增强,包括高阶网络与感觉和运动网络的网络间连接。相反,感觉和运动网络(SMN)的网内rich-club联系明显减弱。与以前研究结果一致,表明丙泊酚引起的连接中断可能发生在低阶RSNs的核心区(例如听觉和视觉网络之间的内部网络连接)。
4、生理性与药物诱导的意识消失的功能网络差异
镇静类似睡眠状态,因为镇静的受试者的大部分回忆都描述其为睡眠。然而,它们是两种独立的无意识状态。总体而言,在丙泊酚作用下,高水平认知网络向感觉和运动网络的rich-club迁移模式与本研究中的自然睡眠模式相似。然而,rich-club区域分布以及丙泊酚诱导的意识消失和自然睡眠之间的联系大有不同。此外,与自然睡眠相比,d-PIS过程中rich-club连接的强度和数量显著增加,而局部连接的强度和数量显著降低。在麻醉药诱导的意识消失期间,我们观察到局部和全部大脑网络的网络效率降低。这些结果表明,与自然睡眠相比,麻醉诱导的意识消失状态下功能性脑网络具有更好的协调性或传递性,但效率较低。
我们也发现丙泊酚诱导和自然睡眠引起意识消失的共同特征。位于双侧角回和枕后回的rich-club两者情况均相同。丙泊酚诱导的意识消失和慢波睡眠都与角回的血流量减少有关。然而位于左侧ACC和双侧IPL和IPS的rich-club仅在丙泊酚诱导的意识消失时出现,而位于右侧额叶上背侧、右侧PCC、左侧顶后回和双侧颞回的rich-club仅在自然睡眠时出现。综上所述,额叶回、中央前回、中岛、PCC和小脑的结节效率可以作为区分麻醉药诱导和睡眠的意识消失的指标。
5、局限性
目前的研究为在系统水平上理解丙泊酚诱导镇静的神经机制提供了一个新的视角,但有着不容忽视的局限性。首先,我们研究的参与者规模相对较小(17名健康志愿者,PIS组8名和睡眠组9名),这可能存在偏倚。个体差异等重要的因素也没有考虑在内。未来的研究需要更大的样本量来支持本研究的结果。其次,由于条件的限制,睡眠状态下的数据采集时间与个体的生物钟不同。需要更严格、更透彻的研究和更好的科学实验设计,以应对其他因素的影响,来验证目前的发现。
五、结论
丙泊酚引起的意识消失使人脑中的rich-club组织发生了广泛的变化,从高阶认知网络(DMN和FPN)到初级感觉皮层(SMN和ON)和小脑网络的rich-club重组。在丙泊酚诱导的意识改变过程中,脑功能网络的整体信息交换增强。生理自然睡眠也表现出类似的rich-club重组模式。rich-club区域从高阶认知网络(DMN)切换到感觉运动网络(SMN,包括部分额叶和颞回)。这些发现表明,大脑网络的rich-club重组可能是生理和药物诱导意识消失的共同的潜在神经机制。此外,与自然睡眠相比,丙泊酚诱导的意识消失过程中,脑岛、后扣带回、额回和小脑等中枢的结节效率显著降低。这表明局部拓扑性质可以作为区分药理性和生理性意识消失的一种手段。未来,中枢重组模式应在其他药物诱导的无意识状态中进行研究,以确定GA的神经机制及其与慢波睡眠的关系。
述评:
全身麻醉是一种药物诱导的、可逆转的昏迷。它由四部分组成:痛觉缺失(analgesia);失去意识(unconsciousness);记忆缺失(amnesia);运动不能(akinesia)。全身麻醉通过保持稳定的血压、心跳和体温使身体处于稳定的状态。常用的麻醉药物与GABA受体结合,这是一种在整个神经系统的抑制性中间神经元中发现的受体类型。这些中间神经元的作用就像计算机网络中的路由器:它们连接并调节大脑中的所有兴奋性神经元。如果你控制了中间神经元,就可以控制大脑中的其他回路。全麻药物和GABA受体的结合激活了这些抑制性中间神经元,从而抑制大脑的点活动。抑制性活动的增强改变了脑电(神经元群体的同步放电)的特征,它们从非常高频、小振幅的波转变为低频、大振幅的波。脑电变得局部化和同步化,无法再传递信息。因此,大脑区域间无法再相互交流,导致深度无意识和记忆缺失。麻醉医师也许会使用阿片类药物(opioid)和肌肉松弛剂,以分别确保充分的镇痛和肌松的状态。当麻醉剂药效逐渐失效时,神经振荡慢慢恢复正常,病人也会清醒过来。2012 年,纪念弗朗西斯•克里克的“意识大会”上宣布的“剑桥宣言”主张自我意识的产生并不依赖特定的大脑构造,实现意识状态重要的是神经复杂性,只要突触的数量足够多,神经系统是什么形状都可以。对睡眠及全身麻醉的研究证明,睡眠、全身麻醉和意识障碍的共同特点是大脑各区域间的功能连接中断、一系列功能状态受到限制,网络效率降低并增加孤立的模块化,为信息传输和集成创造了不适宜的条件。功能性核磁共振,Functional Magnetic Resonance Imagine, 简称fMRI,是医学临床治疗,神经科学研究中一个比较常用的脑成像技术。大脑的血流水平与大脑的神经活跃是相关的。这个血氧含量信号的变化,在英文中叫做blood-oxygen-level-dependent signal, 简称BOLD signal.
而fMRI就是通过对比任务前的BOLD信号与任务中的BOLD信号的差异,来反映大脑活跃的区域及活跃的程度。在分析神经网络的解剖-功能构成时,神经系统被模拟成一系列的节点(解剖和功能的神经元)和内在网络的边(结构-功能连接)。这种脑模型结构有两个基本属性:不同功能的脑区彼此分隔(包括认知、感觉运动整合、辨识和行为);不同功能区通过神经元和区域间连接将功能整合。而神经影像学技术能将脑活动可视化,脑网络分析是探索结构-功能网络的有力工具。目前脑网络研究的一个重要的挑战是找到简单但有意义的方法来描述大脑网络,同时避免任意选择,并从网络数据中提取新的诊断措施或生物标记。Golkowski等研究全身麻醉药物的fMRI发现,全身麻醉状态的网络内神经活动及网络间功能连接均受到抑制,全身麻醉药物对不同脑区活动的改变呈层级性与区域异质性;丙泊酚及七氟醚均能够使大脑活动的动态性降低,网络间连接模式分布改变,利用SVM模型选取网络间连接模式、联合网络间连接及网络内活动为先验特征区分两种状态的准确率显著高于单一网络内神经活动,表明大脑特定的网络连接模式对于区分不同意识状态具有重要作用,特定网络活动的降低是影响网络间功能连接的重要因素。本文将EEG和fMRI结合使用可实现精细的空间分辨率和准确的时间分辨率集成,也是从不同的维度对麻醉和睡眠引起的意识状态的改变进行比较研究。引用中科院智能卓越创新中心徐敏教授访谈中的一段话:脑科学目前还处于起步阶段,包括在神经系统的发育、各种行为的神经基础、神经系统的疾病、意识状态变化深层次的原因等问题上都没有一个明确的答案。其中最根本的原因还是大脑太复杂了,而我们目前的研究手段还相对不足。因此,亟待解决的难题之一就是研究工具的创新。比如,如何在高时空分辨率上大规模记录行为中的神经活动。开发更加高端的技术手段在介观和微观层面分析我们的神经系统,也是未来的挑战。但可以相信在将来随着科技的发展,科学能够更加深入的了解大脑。
原文链接:
Wang S, Li Y, Qiu S, Zhang C, Wang G, Xian J, Li T, He H. Reorganization of rich-clubs in functional brain networks during propofol-induced unconsciousness and natural sleep. Neuroimage Clin. 2020;25:102188. doi: 10.1016/j.nicl.2020.102188. Epub 2020 Jan 21. PMID: 32018124; PMCID: PMC6997627.
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您

















































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




