
北工大:基于代谢组学与群体感应层面的低碳氮比下部分反硝化污泥颗粒化识别(国人佳作)
编译:微科盟HushKuo,编辑:微科盟茗溪、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
部分反硝化颗粒污泥(PDGS)和反硝化颗粒污泥(DGS)在废水脱氮中起着重要作用。然而,二者的聚集能力与COD/氮的比率(COD/N)的内在原因尚未清楚。本研究结合代谢组学分析、微生物分析、颗粒性能和胞外聚合物(EPS) 结构分析等手段,探索不同进水COD/N比下的颗粒污泥形成机制。结果表明,较高的COD/N选择性地增强了糖异生途径、嘌呤和嘧啶代谢途径,导致更多的胞外多糖(PS)释放和絮状污泥生成。碳源的缺失减弱了三羧酸循环(TCA)反应,导致NAD+和ADP减少,亚硝酸盐积累和微生物群落结构发生变化。低COD/N比下氨基酸生物合成途径增强,促进了EPS的疏水性。在运行期间,PDGS比DGS具有更强的基于酰基高丝氨酸内酯(AHL)的群体感应(QS)。CO8-HSL、C8-HSL和C6-HSL作为AHL的主要形式,在DGS和 PDGS中起主导作用。批量实验表明,添加AHL明显改善了氨基酸、苏氨酸(Thr)、色氨酸(Trp)、蛋氨酸(Met)和甘氨酸(Gly)的合成。仅在高COD/N比率下,AHL的添加才能调控PS合成。Glucose-6P、glycerate-3p和UDP-Glc仅在DSG中发生上调,从而增加了EPS中的亲水基团。结果不仅为深入了解反硝化颗粒污泥的代谢提供了新的视角,同时也表明了颗粒污泥启动和运行相关技术的应用潜力。
图文摘要
论文ID
原名:Identification of partial denitrification granulation enhanced by low C/N ratio in the aspect of metabolomics and quorum sensing
译名:基于代谢组学与群体感应层面的低碳氮比下部分反硝化污泥颗粒化识别
期刊:Chemosphere
通讯作者:曾薇
通讯作者单位:北京工业大学城镇污水深度处理与资源化利用技术国家工程实验室
DOI号:10.1016/j.chemosphere.2021.131895
实验设计与方法
结果
1 不同C/N比下硝酸盐去除及亚硝酸盐积累性能
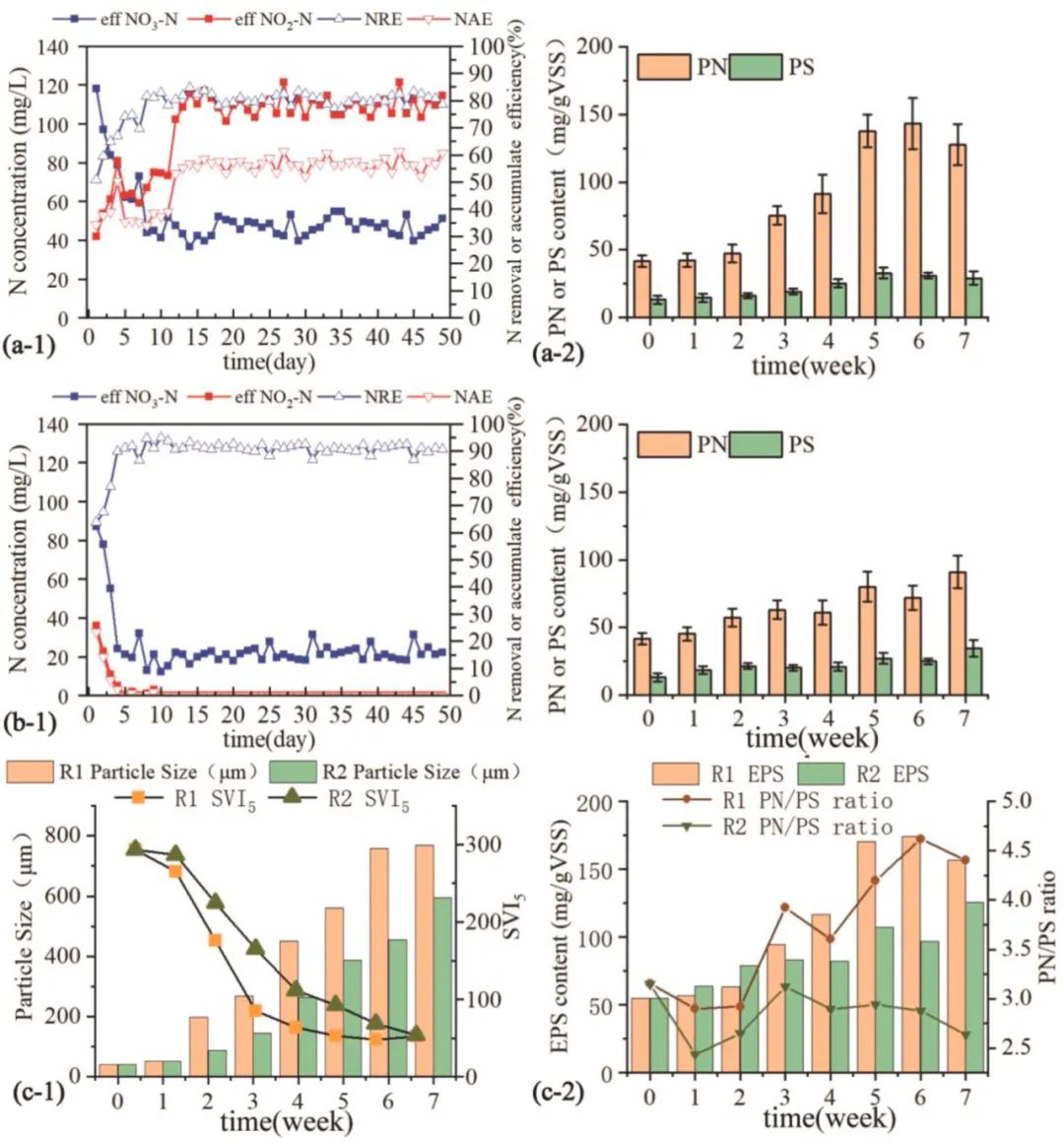
R1和R2中硝酸盐去除和亚硝酸盐积累的性能如图1a-1和b-1所示。R1的不稳定启动时间为12天,比R2(5天)长。在R1运行期间,硝酸盐去除效率(NRE)从50.1%提高到80.5%,亚硝酸盐累积效率(NAE)也从34%提高到53%。R2中NRE也不断加速,第5天达到90.1%。稳定运行后,在低C/N比进水的R1中,观察到NAE为50%~60%,NRE为74.1%~81.5%。在部分反硝化作用下,R1中出水NO2--N浓度达到75-85 mg/L。相比之下,在具有足够碳源的R2中实现了86.5%-92.4%的更高NRE。在完全反硝化的R2中,出水NO2--N浓度几乎为0,出水NO3--N浓度约为20 mg/L。与C/N比较高的R2相比,R1的亚硝酸盐积累量更高,启动时间更长,脱氮率更低,这是由R1中碳源不足造成的。
图1 PDSG和DSG在49天运行期间的性能。出水NO3--N和NO2--N,(a-1)PDSG和(b-1)DSG的硝酸盐去除效率和亚硝酸盐积累效率;(a-2)PDSG和(b-2)DSG的PN和PS组分;(c-1)PDSG和DSG的粒径和SVI5;(c-2)PDSG和DSG中PN和PS的总和和比率。
2 不同C/N比下的反硝化污泥颗粒化
随着粒径的增大和SVI5的减小,颗粒污泥的发育形成过程分为三个阶段(图1c-1、S1和S2):初始阶段(第0-2周的阶段I)、成熟阶段(第2-5周的II期)和稳定期(第 5-7 周的 III 期)。种子污泥的平均粒径、VSS/SS、SVI5和Span值为50.7 μm、0.63、287 mL/g和1.09。培养2周后,污泥絮团聚集成碎片状。在第14天,R1和R2中的SVI5分别降至176 mL/g和225 ml/g,粒径分别升至197.2 μm和136.9 μm。在第二阶段,污泥碎片逐渐堆积成颗粒状污泥。在此阶段(第 14-35天),R1和R2中两个反应器的MLSS分别从4.2 ± 0.6 g/L逐渐提高到7.5 ± 0.3 g/L,以及从4.8 ± 0.2 g/L提高到8.8 ± 0.4 g/L。R1和R2的平均粒径分别增加到560.5 μm和387.8 μm。由于粒径和无机物的增加,R1中的SVI5急剧下降至53 mL/g,而R2中的SVI5为93 mL/g。颗粒在培养5周后成熟,颗粒性质在维持期稳定。在第49天,R1和R2的平均粒径、SVI5和Span值分别为 768 μm和594 μm、52 mL/g和48 mL/g、1.56和2.09。R1中较大的粒径和较小的SVI5表明PDSG在低C/N比进水下具有更好的颗粒性能和沉降能力。R2中较大的Span值意味着DSG的尺寸范围很广,这表明较大和较小颗粒之间存在显著的尺寸差异。相比之下,R1的低Span值意味着PDSG尺寸分布密集(图S2)。
两个反应器中污泥EPS的含量如图1a-2、b-2和c-2所示。从第0周到第5周,R1(从54.8到170.4 mg/g VSS)和R2(从54.8到107.4 mg/g VSS)的EPS 组分(PS和PN)分别增加了115.6和52.6 mg/g VSS。进水COD/N比低的颗粒污泥可以合成更多的EPS,有利于颗粒化。运行结束时,PDSG和DSG中颗粒的PS含量分别为29.1 mg/g VSS和34.5 mg/g VSS。PDSG的PN含量为127 mg/g VSS,高于DSG。在进水COD/N比较低的PDSG中,PN/PS比为4.4,而在DSG中为2.6(图1c-2)。
用FTIR和XPS分析了EPS中的官能团和元素组成。R1中FTIR峰的位置和类型与R2不同,表明EPS中化学基团的差异(图2a-1、a-2和表1)。R1中大约1384 cm-1处的峰值水平明显高于R2(在1363 cm-1处),这是与-CH3相关的胺和脂类中的C-H延伸。1449 cm-1处的谱带是甲基的不对称角峰,该峰可在R2的EPS中明显观察到,但在R1中不可见。大约1649 cm-1处的条带归因于与蛋白质相关的C=O(C=C)拉伸,此现象在R2中不可见,仅出现在R1中。
研究用FTIR波段分析了蛋白质二级结构(图2b-1和b-2)。图2(d)分别显示了R1和R2的EPS中的四种蛋白质二级结构:α-螺旋、β-折叠、无规卷曲和β-转角。R1颗粒EPS中的蛋白质含有37.2%的β-折叠、24.2%的α-螺旋、20.6%的无规卷曲和19.8%的β-转角,R2具有24.2%的β-折叠、26.4%的α-螺旋、25.2%的无规卷曲和19.8%的β-转角。因此,α-螺旋/(β-折叠+无规卷曲)的值在R1中为0.389,在R2中为0.535。如果α-螺旋含量低而β-折叠和无规卷曲含量高,则PN结构松散,导致内部疏水位点暴露,颗粒沉降性能良好。
通过X射线光电子能谱测试显示EPS中的元素结合能,描述元素形式。图2c-1和c-2显示了R1和R2中的C-1s XPS光谱。研究定量了EPS中C-C/H、C-OH、C=O、O=C-O和C-O-C的五种主要的C元素形式(图2d)。EPS的主要疏水成分C-(C/H)的形式在R1中占51.4%,比R2中高8.2%。此外,R1 EPS的其他亲水元素形式(C-OH、C=O、O=C-O)仅占46%(15.0%、14.3%和16.7%之和),明显低于R2中的55.6%(18.8%、9.2%和27.6%的总和)。XPS结果进一步证明PDGS在EPS中含有更多的疏水成分。
图2 R1和R2中EPS的化学结构。PDSG EPS(a-1)和 DSG EPS(a-2)的FTIR;PDSG EPS(b-1)和DSG EPS(b-2)的蛋白质二级结构;PDSG EPS(c-1)和DSG EPS(c-2)的XPS;PDSG和DSG EPS的蛋白质二级结构和元素C形式的值(d)。
表1 R1和R2中EPS的类型和位置。
3 响应不同C/N比的代谢物谱
在运行期间的最后一天,对GS样品进行了基于LC-MS的代谢组学分析和定量分析。代谢组学一直被用于例如肠道菌群和发酵系统等混合微生物系统的研究。研究检测到了51种代谢物,火山图中显示了PDSG和DSG之间的不同代谢物(图3a)。结果表明,由于C/N比的不同,20多种代谢物的含量发生了显著变化。如图3b 所示,从代谢物浓度获得基于PCA评分的散点图显示了两组之间的明显分离,表明进水COD/N比的变化对GS代谢途径有显著影响。相关代谢物和代谢途径如图3c所示。PDSG和DSG的代谢产物在能量过程、氨基酸和多糖合成途径、电子传递等方面存在显著差异。三羧酸循环(TCA)中间体(如富马酸、苹果酸、柠檬酸、乌头酸和琥珀酸)的相对含量在进水COD/N比较低的PDSG中迅速下降。此外,DSG中3-磷酸甘油酸(Gly-3-p)、6-磷酸葡萄糖(Glc-6-p)、葡萄糖(Glc)、果糖(Fru)和海藻糖高于PDSG,这意味着在较高的进水COD/N比下,DSG中的糖异生途径更活跃。一些类核苷酸代谢物(如鸟嘌呤、胞嘧啶、硫胺素和腺嘌呤)在细菌生长中发挥重要作用,这些代谢物在DSG中被上调。研究共检测了出14种氨基酸,包括谷氨酸(Glu)、精氨酸(Arg)、脯氨酸(Pro)、赖氨酸(Lys)、蛋氨酸(Met)、组氨酸(His)、苏氨酸(Thr)、亮氨酸(Leu)、缬氨酸(Val)、丙氨酸(Ala)、酪氨酸(Tyr)、甘氨酸(Gly)、天冬氨酸(Asp)和丝氨酸(Ser)。Gly、Met、Pro、Tyr和Leu在PDSG中增加,表明在低进水C/N比下,PN合成途径在PDSG中更加活跃。
图3 从PDSG和DSG样品中提取的代谢物的统计分析。(a)火山图,红点代表组间差异显著的代谢物;(b)PCA分析,不同COD/N比下的样品聚集在一起。(c)与DSG相比,PDSG相关途径的代谢组学变化。圆圈的颜色代表PDSG与DSG代谢物的相对丰度比。
4 不同C/N比下的AHLs变化
研究每周测定AHLs以描述整个运行期间的QS。在QS群中确定了三个具有代表性的AHL(C6-HSL、C8-HSL和CO8-HSL)。颗粒形成过程中三种AHLs 含量的升高,表明QS随着絮状污泥的聚集而增强(图4)。开始时,两个反应器含有单一类型的C8-HSL,少量为0.07 μg/g VSS。然而,随着颗粒的形成,AHLs的含量显著增加。从2到5周,R1中C6-HSL、C8-HSL和CO8-HSL的浓度从0.36 ± 0.1、0.35 ± 0.08、0.16 ± 0.04 μg/g VSS分别增加到0.53 ± 0.16、0.77 ± 0.25、0.27 ± 0.11 μg/g VSS。与R1相比,R2中AHLs的含量从0.21 ± 0.07(C6-HSL)、0.28 ± 0.09(C8-HSL)和0.17 ± 0.06(CO8-HSL)μg/g VSS增加到0.42 ± 0.18、0.44 ± 0.11、0.21 ± 0.09 μg/g VSS,但小于R1。此后,AHLs在第5周到第7周保持相对稳定。简而言之,AHLs介导的QS在颗粒化过程中明显增强,并且在PDSG中达到比DSG更高的含量。AHLs的含量和变化趋势与其他研究颗粒污泥系统中的相似。
图4 R1和R2中的AHL浓度变化。RiWj表示第j周来自反应器i的样品。
5 不同C/N比下的AHLs调控
为了探索GS性能与AHL差异的内在机制,共进行了24次批次检测(2种污泥×4 AHL条件×3个平行实验),每个样品检测到51种代谢物。单向方差分析表明对照组和测试组之间代谢物具有明显差异(图5a-1和b-1)。从代谢物浓度获得的基于PCA评分散点图,显示了不同C/N比下(PDSG组和DSG组)四个样品(对照、添加C6-HSL、添加C8-HSL和添加CO8-HSL)的明显分离(图5a-2和b-2)。每个样品中均能观察到响应不同AHL的代谢池的明显变化。相关的代谢物和代谢途径见图6。同种AHLs不同进水C/N比引起不同的代谢变化,说明C/N比是影响AHLs调控的重要因素。C8-HSL和CO8-HSL增加了甘油酸(Gly-3-p)、尿苷二磷酸葡萄糖(UDP-Glc)、葡萄糖(Glc)和海藻糖的合成,然而在加入C6-HSL和C8-HSL的PDSG中不明显。此外,添加任何AHL后,GS代谢物中苏氨酸(Thr)、色氨酸(Trp)、蛋氨酸(Met)和甘氨酸(Gly)的含量均增加。
图5 从添加AHL测试样品中提取的代谢物的统计分析。PDSG(a-1)和DSG(b-1)的单向方差分析。红点代表四组差异显著的代谢物;(b)PDSG(a-2)和DSG(b-2)中的PCA分析。“空白”、“C6”、“C8”和“CO8”分别代表对照、添加C6-HSL、C8-HSL和CO8-HSL。
图6 II期AHL调控的代谢物和代谢途径。描述了(a)中心碳代谢、(b)糖异生途径和(c)氨基酸生物合成途径中的碳水化合物代谢。热图中显示了添加C6-HSL、C8-HSL、3OC8-HSL的对照和GS中代表性代谢物谱。标记了外源性AHL显著富集的代谢物。
讨论
1 代谢输出的差异改善部分反硝化
代谢组学已成为揭示微生物代谢和调控机制的有效方法。TCA循环作为反硝化细菌的能量中心,为反硝化反应提供还原动力。NAD+和NADH是TCA循环和反硝化反应之间的重要电子载体,它们在微生物代谢活动中起着重要作用。在PDSG中,NAD+的相对含量(图3c)随着TCA反应的减弱而降低,不能为完全反硝化提供足够的还原活性。硝酸盐还原酶比亚硝酸盐还原酶具有更强的电子亲合力,因此在电子供体有限的低COD/N流入物下,硝酸盐是优选的电子受体。对于Thauera spp.,当硝酸盐存在时,亚硝酸盐还原酶基因转录受到抑制,由于NAD和TCA途径水平较低,这样可以节省大量能量并带来竞争优势。在这项研究中,Thauera spp.的丰度在低进水COD/N比率下显著增加(图S3)。这些证据表明,上调或下调途径是由进水COD/N比值引起的,影响了微生物群落结构并促进了亚硝酸盐的积累。
2 细菌聚集能力代谢增加差异
在核苷酸碱基通路中发挥重要作用的代谢物,如鸟嘌呤、胞嘧啶和硫胺素,在DSG中得到了上调(图3)。这些代谢物参与多种代谢过程,包括蛋白质合成、DNA合成和细胞稳定,促进细胞增殖和微生物生长。在本研究中,参与细胞物质合成的嘌呤和嘧啶代谢途径上调,导致MLSS和MLVSS增加(图S1)。与颗粒污泥相比,絮状污泥具有更大的比表面积和更强的代谢活性,导致絮状污泥含量和Span值增加(图S2)。由此可见,随着反应器运行时间的延长,颗粒污泥的平均粒径减小,SVI5升高,影响了GS系统的长期稳定运行。这些证据表明,上调或下调的途径影响了污泥的形式。
3 基于AHLs的QS和调控促进低C/N比下的污泥颗粒化
颗粒污泥系统中AHLs的浓度随着污泥粒径的增加而增加(图1c-1,图4)。研究人员发现,较高的AHL是在不合适的情况下产生的,例如营养过剩/饥饿、波动的HRT、波动的C/N比或高盐度。大多数反硝化细菌都含有与AHLs合酶相关的基因,如LuxI和HdtS。实验结果表明,部分反硝化微生物群落Thauera spp.(R1中63.4%,R2中36.5)在低C/N比条件下提高了QS并形成颗粒,减少了缺乏碳源的负面影响。此外,通过在碱性pH下破坏内酯环,AHL很容易被降解,导致QS信号的缺失。在部分反硝化过程中,NO3-被还原为NO2-而不是N2,与完全脱氮过程中相比,产生的OH-更少。AHLs免受液相中过量碱的影响,由于R1中的部分反硝化作用,QS得到了增强。
在AHLs回加测试中,所有AHLs都上调了氨基酸的水平,例如Thr、Trp、Met和Gly(图6)。Asp和Leu等氨基酸对于细胞外PN合成和生物膜稳定是必不可少的。之前的研究表明,细胞中Ala、Val和Gly等氨基酸的浓度与其在EPS中的浓度密切相关。此外,8碳侧链AHL与EPS中类色氨酸物质的浓度呈正相关。因此,AHLs主要通过上调氨基酸合成途径促进PN的产生,尤其是Thr、Trp、Met和Gly的产生,导致了絮体大小的增加(图1c-1和6)。
根据AHLs回加测试,外源性AHLs仅在DGS中增强糖异生途径(图6),这是多糖合成的基本途径,决定了EPS中PS的含量。较低的PN/PS比有利于污泥的聚集能力和疏水性,有利于颗粒化过程。相比之下,在外源性AHLs加入PDSG的情况下,糖异生途径中的大部分中间体,如Gly-3-p和UDP-Glc没有明显变化。然而,PDSG和DSG中的优势细菌Thauera spp.具有完整的糖异生途径,表明进水COD/N改变了QS的调节途径。在PDSG中,乙酸不足会降低草酰乙酸和Glc-6-p的含量,削弱AHLs对糖异生途径的调控。在以往的研究中,在AHLs处理下,PS的增加因环境和进水而异,变化范围在1%-40%之间。
4 低C/N比导致疏水性EPS有利于污泥颗粒化
以乙酰辅酶A开始的TCA循环是进水乙酸盐的活化方式,是生物合成的重要代谢途径。如图3所示,在PDSG进水乙酸盐含量较低的情况下,TCA循环中间体的相对含量迅速下降,导致PS生物合成的底物缺乏。在这种情况下,由于以下原因,PN生物合成占主导地位。首先,微生物可以通过多种途径合成氨基酸,但制造多糖前体的途径只有一种。其次,不同种类的氨基酸可以很容易地相互转化。最后,许多多糖的前体用于氨基酸合成,例如Asp(由草酰乙酸合成)、Val和Leu(由丙酮酸合成)、Ser(由磷酸烯醇丙酮酸合成)。氨基酸合成的优势增加了胞外蛋白的相对含量,从而提高了PDSG的EPS 中δ C-H链和C=C的含量。XPS分析还表明PDGS中的EPS含有更多疏水性官能团(C-C/H)以及强化的氨基酸合成途径(图2d)。EPS中的PN与活性污泥的表面疏水性特别相关,其影响细菌表面电荷、疏水性和聚集体空间结构。这证明了PDGS的表面疏水性比DGS更强,并证明细胞内代谢的变化与细胞外化学基团有关。与DSG相比,PDSG中EPS的二级结构具有较少的α-螺旋和更多的β-折叠,这决定了PN的疏水性。当α-螺旋含量远大于β-折叠和无规卷曲含量的总和时,PN具有松散结构。这种松散的PN结构导致内部疏水位点的暴露,提高了EPS的疏水性。如图3所示,PDSG中疏水性氨基酸的含量高于DSG,尤其是Tyr和Met。这从氨基酸组成分析的角度暗示了EPS中蛋白质更亲水的特性。疏水氨基酸R基团的特性直接影响蛋白质的表面性质,因此疏水氨基酸的比例影响蛋白质的表面疏水性。EPS的强疏水性有利于污泥的聚集,这与DSG和PDSG的颗粒化一致。
EPS的疏水性受进水和微生物群落的影响。以往的研究发现,颗粒污泥在进水C/N比较低的条件下变得稳定,从而提高了EPS的PN含量和疏水性。Thauera属具有EPS分泌功能,在C/N比低于4时出现,在污泥颗粒化中起关键作用。EPS的疏水成分对微生物聚集和絮凝能力具有显著影响,与颗粒稳定性直接相关,这解释了部分反硝化污泥颗粒化的原因。
结论
与完全反硝化颗粒污泥相比,部分反硝化颗粒污泥在低COD/N进水下具有更强的基于AHL的群体感应、更多的PN含量和更好的沉降性能。3种AHL(C6-HSL、C8-HSL和CO8-HSL)在运行期间有所增加。AHLs选择性地促进氨基酸的合成,如Met、Thr、Gly、Trp和Met,促进细胞外蛋白质的含量。较高的COD/N比率选择性地增强了糖异生途径,导致更多的细胞外PS排泄。较高的COD/N比值也增加了类核苷酸代谢物的含量,导致絮泥和Span值的增加。碳源缺乏削弱了TCA循环,导致NAD+减少、亚硝酸盐积累和微生物群落结构发生变化。对PDSG和DSG微生物之间代谢差异的认识为揭示颗粒化中的代谢机制提供了新的视角,同时进一步论证了与颗粒污泥启动和运行相关技术的潜在应用。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您








































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




