
尽管取得了这些相对迅速的进展,但我们对肠道病毒特性的理解仍然存在重大差距。
编译:微科盟小木,编辑:微科盟茗溪、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
肠道微生物组包括一系列微生物基因组,如细菌组、病毒组、真菌组等。肠道微生物群在肠道免疫和疾病中起着至关重要的作用,例如炎症性肠病(IBD)和结直肠癌(CRC);然而,其潜在的机制仍未被完全理解。阐明微生物群和炎症之间的关系可能会极大地提高我们对病因、疾病进展、患者管理以及预防和治疗发展的理解。在这篇综述中,我们讨论了肠道病毒(即共生病毒、致病性病毒和噬菌体)在结肠炎和结直肠癌的发生、发展和并发症中的影响,以及它们在预防和治疗方面的潜力。我们探讨了肠道病毒和宿主免疫系统之间的相互作用对炎症疾病和癌症的严重程度的影响,包括肠道病毒与其他微生物和微生物产物之间的直接和间接相互作用。此外,通过具体临床试验的例子,解释了病毒在肠道炎症反应中的作用的潜在机制,以推断潜在的治疗靶点。鉴于迄今为止讨论肠道病毒群的不同主题的文献非常有限,我们相信这些广泛的分析可能有助于深入了解IBD和CRC的分子发病机制,这可能可以帮助改进这些重要的人类疾病的治疗方案。
论文ID
原名:Dynamic impact of virome on colitis and colorectal cancer: immunity, inflammation, prevention and treatment
译名:病毒组对结肠炎和结直肠癌的动态影响:免疫、炎症、预防和治疗
期刊:Seminars in Cancer Biology
IF:15.707
发表时间:2021.10.14
通讯作者:黄灿华&Min Wu
通讯作者单位:华西医院生物治疗国家重点实验室、肿瘤中心,四川大学华西基础医学与法医学院,成都生物治疗协同创新中心;美国北达科他州立大学医学与健康科学学院
DOI号:10.1016/j.semcancer.2021.10.004
综述目录
1 前言
2 病毒在人体肠道中的分布和丰度
3 与肠道炎症和癌症密切相关的病毒
4 可能与肠道炎症和癌症有关的病毒
5 以噬菌体为基础的肠道炎症和癌症治疗
6 治疗CRC的溶瘤病毒
7 研究肠道病毒群的新兴技术
8 挑战与未来研究方向
附:缩写
主要内容
根据国际癌症研究机构(IARC)发布的GLOBOCAN 2020,结直肠癌(colorectal cancer, CRC)是2020年世界上第三大常见癌症(发病率10%)和癌症死亡的第二大原因(死亡率9.4%)。既往研究表明,慢性炎症与结直肠癌的病因和发病机制有关,称为结肠炎相关CRC(CAC)。炎症性肠病(Inflammatory bowel disease, IBD),包括克罗恩病(Crohn’s disease, CD)和溃疡性结肠炎(ulcerative colitis, UC),显著增加了CAC的风险,例如,UC患者发生CAC的风险增加了2.4倍。CD患者的CAC发病率相似,但年龄低于健康人群。尽管CRC的分子机制尚不清楚,但一种可能性是与遗传易感个体对肠道微生物群的免疫反应加重有关。反过来,微生物群也可能显著影响宿主防御和炎症反应,从而影响IBD和CRC的发展和进展。
肠道微生物群是一个复杂的生态系统,在人类健康和多种疾病的发病机制中发挥着重要作用。近几十年来,随着高通量、新一代测序技术和生物信息学的进步,数万亿细菌、真菌、病毒、寄生虫和古生菌已被确认。已发现细菌(胃癌的幽门螺杆菌)和病毒(肝癌的乙肝和丙肝病毒以及宫颈癌的人乳头瘤病毒)强烈影响癌症发生(肿瘤发生)、进展和治疗。结肠炎和癌症微生物组研究几乎完全集中在细菌成员基因组上。目前,在估计的1012种不同的微生物中,只有11种被认为是人类致癌物。越来越多的证据表明,仅靠某些微生物的存在和数量改变不足以致癌,但它们可以作为“第二击”,促进致病性细菌感染、生态失调和宿主免疫调节的遗传缺陷引起的炎症和癌变。目前,大规模的临床试验正在测试微生物群对癌症治疗的影响,包括诊断、饮食调整和肿瘤内注射工程细菌。
几十年的研究只发现了几种直接导致癌症的病毒,但似乎有很多病毒对宿主的免疫系统有复杂的影响。越来越多地认识到病毒感染参与IBD或CRC的情况。CRC是一个循序渐进的过程。病毒稳态失调可能引发或促进炎症性疾病(如IBD),促进发育不良,最终导致症状严重和高死亡率的癌症。已有研究表明,病毒直接参与炎症和肿瘤的发生,因为它们有感染人类细胞和突变的能力。此外,病毒可以通过调节细菌群落的稳定性和组成间接起作用。因此,病毒被认为是潜在的模块化生物疗法。进一步了解病毒如何影响IBD和CRC,对于早期发现和预防癌症早期阶段之后的进展有很大的潜力,并显著促进治疗方法的发展。
在本文中,我们回顾了目前将病毒群与炎症反应、结肠炎、CRC的发生和进展联系起来的知识。本文总结了黏膜病毒的组成及其在健康和疾病中的直接作用(如结肠炎和CRC)。此外,本文还讨论了病毒通过调节相关的细菌群落间接影响结肠炎和癌症。最后,我们尝试填补知识空白,并指出未来潜在的研究方向。
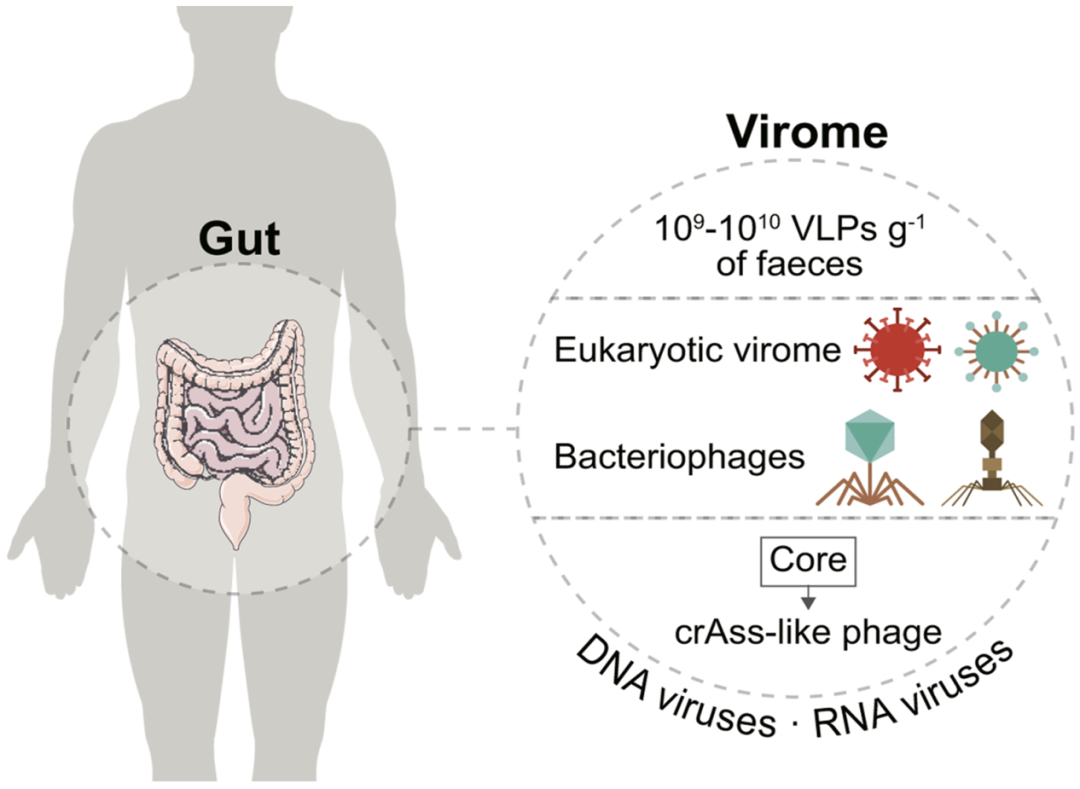
人类肠道病毒群包含肠道中自然存在的所有DNA和RNA病毒的基因组,包括噬菌体、真核病毒、古病毒和内源性逆转录病毒(图1)。以往的研究主要集中在细菌(噬菌体)和真核病毒。除此之外,由于对肠道病毒群在人类中的作用了解有限,且病毒数据库相对较小,所以对肠道病毒群的其他组成部分的研究还不够深入。
噬菌体(phages)是细菌病毒中种类最丰富的一类,它通过免疫调节和杀菌作用来影响体内稳态。2003年首次公布了从单个健康个体中分离出来的人类肠道病毒群,每克至少有109-1010个病毒样颗粒(virus-like particles, VLPs)(图1)。肠道病毒群包括近1015个噬菌体,比细菌细胞多10倍,比人类细胞多100倍。以双链DNA(dsDNA)噬菌体有尾病毒目(Caudovirales)和单链DNA(ssDNA)噬菌体微病毒科(Microviridae)为主的群居病毒具有高度的特异性、多样性和稳定性。然而,这些噬菌体中的大多数仍未分类。核心噬菌体群落,包括crAss样噬菌体,是人类肠道中最丰富和最广泛的病毒(20-50%)。与非洲群体相比,西方社会crAss样噬菌体的组成有显著差异。此外,最近在孟加拉国和坦桑尼亚Laksam地区的个体中发现了一种独特的流行噬菌体分类群(LAK噬菌体)。由于共生病毒参与宿主免疫培养和成熟,这些病毒的生态失调可能会导致IBD和癌症,我们将在这篇综述的后面进行讨论。
图1 病毒在人体肠道中的分布。人类病毒群包含DNA和RNA两种类型,人类每克毛重粪便中含有109-1010个病毒样颗粒。病毒群的主要成分是真核生物、细菌(细菌噬菌体或噬菌体)、古菌病毒和内源性逆转录病毒。核心噬菌体群落是人类肠道中最常见的噬菌体,例如crAss样噬菌体。该插图是通过改编SMART (https://smart.servier.com)和Vecteezy(vecteezy.com)模板创建的。
多种肠道病毒可以感染人类,包括逆转录酶病毒、诺如病毒(norovirus)、轮状病毒、腺病毒和疱疹病毒。例如,诺如病毒是全世界范围内食源性肠胃炎的主要病因。小鼠诺如病毒(MNoV)在小鼠体内形成终身肠道感染,无症状个体可长期携带诺如病毒而无明显疾病。肠道病毒感染可导致从无症状到中度、重度急性或重度慢性疾病的病理生理过程,可能会导致结肠炎和癌症(图2)。
在过去的20年里,随着病毒宏基因组学的发展,病毒与肠道炎症和癌症之间的研究领域已经从检测病毒粒子的存在发展到相互作用和病毒驱动的分子机制的研究(表1)。
图2 宿主-病毒在肠道炎症和癌症中的相互作用。肠道病毒丰富度随着IBD和CRC患者病情的加重而增加,而细菌多样性和丰富度则降低,反映出疾病与细菌和病毒微生物的反向相关性。肠道炎症的诱导或直接的遗传毒性作用是微生物群诱导肠道癌变的主要机制。病毒可通过①直接感染肠道;②通过细菌间接影响宿主;③与宿主免疫系统相互作用并诱导免疫应答等途径促进CRC的发生。该插图是通过改编SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板创建的。
表1 病毒与肠道炎症和癌症的关系。
IBD患者的噬菌体组成与健康对照明显不同。与微病毒科(Microviridae)相比,CD或UC患者的有尾病毒目(Caudovirales)显著增加,细菌丰富度和多样性降低。在儿童CD患者的回肠活检中也发现Caudovirales增多,但粪便中Caudovirales增多与UC发病无关。在另一项研究中,发现结肠中有疱疹病毒科(Herpesviridae)序列的患者显示出人类内源性病毒基因表达增加和微生物多样性增加。在早期诊断的未接受治疗的IBD患者中,UC患者的Hepadnaviridae转录本数量高于CD患者和对照组,而Polydnaviridae和Tymoviridae含量较低。此外,与对照组相比,CD患者的Hepeviridae丰度增加,Virgaviridae丰度减少。在IBD患者中普氏栖粪杆菌(Faecalibacterium prausnitzii)通常减少,在IBD患者的样本中发现F. prausnitzii噬菌体明显高于家庭对照组,表明IBD中F. prausnitzii介导的死亡率增加。值得注意的是,特异性病毒感染可与IBD风险基因相互作用,改变IL-10或Atg16L1缺陷小鼠的肠道疾病,这表明某些种类的病毒可能会导致IBD的发生(图3)。
图3 肠道病毒对肠上皮细胞和宿主免疫细胞的潜在影响。在IL-10缺陷小鼠中,MNoV通过①微皱褶细胞穿过上皮屏障,②感染免疫细胞(如淋巴细胞、巨噬细胞和树突状细胞),并③引起干扰素反应(IFN-α,β)。在IL-10缺陷小鼠中,MNoV可④感染簇状细胞,⑤诱导IFN-λ分泌,其对体内病毒的持续调控起关键作用。⑥持续性MNoV也可导致与IBD相关的Paneth细胞异常。⑦MNoV感染Atg16L1突变小鼠引发Paneth细胞异常,表现出肠道疾病。IBD, 炎症性肠病; IEC, 肠上皮细胞; MNoV, 小鼠诺如病毒。该插图是通过改编SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板创建的。
CRC患者和健康对照组之间的病毒丰富度没有显著差异。然而,据报道,病毒失调与早期和晚期CRC有关。虽然病毒在CRC发病机制中的影响越来越大,但至今还没有明确一致的共识结论。进一步严格的实验和交叉队列验证是解决这一谜题的关键,建立或反证病毒与CRC的联系,特别是在临床患病率和病理生理学上。
在CRC标本中检测到人乳头瘤病毒(human papillomavirus, HPV)基因组,表明HPV的存在可能与患CRC的风险增加有关。然而,其他研究质疑HPV在结直肠癌发生中的作用,因为在结直肠癌标本中很少或没有发现HPV DNA,这强调需要进一步的研究来阐明这种不一致。与HPV阴性组织相比,四种差异表达基因(WNT-5A, c-Myc, MMP-7和AXIN2)在HPV阳性CRC样本中上调,这与一项早期的报告表明HPV与CRC发病机制相关一致。
有研究表明,人类巨细胞病毒(cytomegalovirus, CMV)优先感染CRC病变组织而非正常健康组织,这可能与CRC患者预后不良有关。这种现象可能与CRC细胞的增殖和进展有关,与对照组相比,CMV感染的CRC样本中TLR2、TLR4、NF-κB、TNF-α表达升高,癌细胞系中Bcl-2、cox-2、Wnt/β-catenin表达升高。
多瘤病毒科(Polyomaviridae)的成员,主要是人类多瘤病毒2(称为John Cunningham virus, JCV)被发现与CRC有关,表明JCV可能具有致癌作用。然而,在一些研究中,CRC样本中很少或没有检测到JCV DNA。有人提出JCV通过以下方式促进结肠癌发生:首先,早期JCV蛋白T抗原(T-ag)被认为介导了病毒的致癌潜能,并与染色体不稳定性有关;其次,JCV可能负责诱导肿瘤抑制基因(如p53)的多态性和/或改变;最后,JCV可能会改变细胞行为(如迁移和侵袭),强调可能参与了PI3K/AKT、MAPK和/或Wnt/β-catenin通路。
在一项罕见的针对意大利人群的研究中,患有乙肝和丙肝病毒相关肝病的CRC患者表现出更好的5年无病生存期和较低的异时性肝转移发生率,作者声称是“金属蛋白酶抑制剂”假说,而不是病毒感染的直接影响。在另一项研究中,发现乙肝病毒(HBV)感染与结直肠癌风险增加密切相关,还有研究发现丙型肝炎病毒(HCV)感染与结直肠癌的发生没有相关性。由于差异显著,有必要进一步研究不同类型肝炎病毒、炎症性疾病、CRC之间的相互关系。
相同的噬菌体在肠道炎症环境中可能扮演着“双刃剑”的角色。有尾病毒目噬菌体可显著减少致癌性细菌的定植,提高易发生CRC的动物的存活率。同样,CD患者中有尾病毒目噬菌体的丰度显著增加。有必要分析这些复杂的相互作用,以测试病毒微生物是否可能在未来的研究中有治疗益处。
因此,尽管有争议,但这一领域的研究已经发现,一些真核病毒可以感染人类细胞,造成感染,触发免疫反应,有时还会导致严重的疾病。未来的研究应探讨病毒组成对特定肠道疾病(如IBD和CRC)的改变和影响。
肠道病毒首先暴露在细菌中,在宿主细胞(肠细胞或免疫细胞)中开始复制。动物感染模型已被普遍用于检查肠道病毒和其他微生物之间的相互作用。目前,这些实验已经通过使用抗生素处理和无菌小鼠或免疫缺陷小鼠和/或感染人类和小鼠病毒的年轻小鼠中进行。细菌微生物群对肠道病毒的有益作用直到最近才被认识到,这可能是因为许多研究涉及腹腔注射病毒而不是自然的口服途径。
由于噬菌体和细菌在进化过程中进行着激烈的“军备竞赛”,噬菌体可能会改变细菌的微生物组,并通过复杂的机制在肠道生理和疾病中发挥作用,这需要进一步广泛的阐明(图4A)。首先,肠道噬菌体负责细菌群落之间的水平基因转移(horizontal gene transfer, HGT),包括发病机制和抗生素耐药性,这给控制细菌感染带来了医疗负担。其次,噬菌体的激活导致其细菌宿主的裂解和特定肠道细菌种类的丰度的变化。最后(但并非最不重要的),细菌裂解会释放作为病原体相关分子模式(PAMP)和抗原的蛋白质、脂类和核酸,这可能触发炎症信号级联,从而诱导细胞因子、细胞浸润和组织损伤。最近的研究表明,病毒可以通过以下机制增强复制和传播(图4B):(1)细菌对病毒粒子的稳定作用。脊髓灰质炎病毒(Poliovirus, PV)通过粪口途径传播,可传播至中枢神经系统。肠道细菌稳定病毒粒子并限制热失活以增强PV复制和小鼠粪-口传播。(2)细菌可增加宿主细胞附着。PV与脂多糖(LPS,革兰氏阴性细菌表面的一种聚糖)和肽聚糖(革兰氏阳性细菌细胞壁的主要成分)结合,增强病毒对宿主细胞的附着。(3)病毒结合LPS可诱导免疫耐受。小鼠乳腺肿瘤病毒(MMTV)是逆转录病毒科(Retroviridae)的一种,在人类中通过母乳从母亲传播给后代。MMTV与LPS结合,诱导宿主TLR信号和IL-10介导的免疫耐受,启动病毒复制和传播。(4)宿主IFN-λ可能受微生物调控。MNoV是杯状病毒科(Caliciviridae)诺如病毒属的一种,通过粪口途径传播,细菌可能通过调节IFN-λ反应促进MNoV的复制。
除了通过细菌间接影响宿主外,噬菌体还可以直接与宿主的免疫系统相互作用,引发免疫反应。越来越多的证据表明,在IBD或CRC患者肠道中,噬菌体与宿主细胞内免疫途径相互作用并激活免疫应答。
共生病毒通过RIG-I-MAVS-IRF1-IL15轴和TLR3和/或TLR7 IFN-β通路抑制上皮内淋巴细胞,保护宿主动物免受右旋糖酐硫酸钠(DSS)诱导的结肠炎(图4C)。有尾病毒目噬菌体间接或直接刺激免疫反应并加重结肠炎。首先,噬菌体裂解细菌并释放促炎产物;其次,噬菌体可以通过TLR9/MyD88依赖的方式促进淋巴结中CD8+和IFN-γ产生的CD4+ T细胞群的增加(图4C)。被金黄色葡萄球菌(Staphylococcus aureus)或铜绿假单胞菌(Pseudomonas aeruginosa)噬菌体感染后,外周血单核细胞表现出转录反应,IL-1、IL-6、TNF的转录明显增强。铜绿假单胞菌感染的人类伤口中的丝状噬菌体触发了免疫细胞中TLR3的激活和I型IFN的产生。反过来,这种类型的细胞因子(IFNα和IFNβ)抑制巨噬细胞产生TNF,从而损害吞噬作用和细菌清除,延迟伤口愈合。尽管在理解噬菌体与细菌的相互作用方面取得了这些进展,但要理解这种“军备竞赛”还为时过早。未来,更多的研究可能会集中在肠道噬菌体对影响噬菌体适应性和发病机制的细菌微生物群组成的影响上。
总的来说,病毒、细菌和宿主免疫系统之间的相互作用正在被更广泛地研究。然而,在许多情况下,因果关系和分子机制仍然没有被完全理解,应该从更广泛的机制上进行探索。
图4 跨界交互作用和机制。A,噬菌体通过宿主相关细菌直接或间接与宿主相互作用。这些相互作用可能影响宿主的遗传变异,并对宿主健康产生明显的影响。B,细菌增强肠道病毒复制和传播的机制。C,病毒影响肠道的可能机制。①树突状细胞可通过RIG-I MAVS IRF1途径感知病毒,刺激IL-15分泌,从而促进IELs增殖,抑制其凋亡。②驻留病毒被树突状细胞TLR3和TLR7识别,产生保护性IFN-β来抑制肠道炎症。③噬菌体被内吞于树突状细胞中,激活B细胞和T细胞,刺激IFN-γ介导的TLR9依赖性免疫反应,从而加剧结肠炎。IEC, 肠上皮细胞; LPS, 脂多糖。该插图是通过改编SMART(https://smart.servier.com)和Vecteezy(vecteezy.com)模板创建的。
以噬菌体为基础的疗法(也称为噬菌体疗法)已经存在了大约100年。60年前,传染病的治疗方法在大多数西方国家几乎被抛弃,因为其结果难以预测,而且新发现的抗生素极大地改善了治疗方法。然而,近几十年来,抗生素的滥用导致了抗生素耐药性的激增。由于缺乏有效的治疗方法和细菌对抗生素耐药性的迅速演变,噬菌体疗法作为多药耐药细菌感染的替代方法重新受到人们的关注,主要是在非常严重的情况下。因此,越来越多的临床报道和研究使用噬菌体治疗致命性细菌感染或其他合并症,如囊性纤维化(CF)和慢性阻塞性肺疾病(COPD)。
这并不奇怪,由于某些致病菌与IBD和CRC都有关联,一些噬菌体相关疗法的临床试验正在进行中,用于治疗结肠炎和CRC。噬菌体治疗可能在微生物操作方面具有优势,因为噬菌体具有针对单一细菌的高度特异性。例如,噬菌体正在针对UC中的艰难梭状芽胞杆菌(Clostridioides difficile)、CD中的粘附侵袭性大肠杆菌(Adherent-invasive E. coli, AIEC)和结直肠癌中的具核梭杆菌(Fusobacterium nucleatum)进行试验。
粘附侵袭性大肠杆菌(AIEC)可能在CD发病机制中起一定的作用。Galtier等人从废水中分离出三种针对AIEC的噬菌体,可以减少AIEC在回肠和结肠定植,减轻DSS诱导的小鼠结肠炎症状。这项工作为CD患者提供了一种新的治疗选择。一项2期双盲、随机、安慰剂对照临床试验(NCT03808103)招募了30例CD患者,以评估AIEC特异性噬菌体鸡尾酒(EcoActive)对疾病活动、炎症标志物和AIEC负荷的有效性。
具核梭杆菌(Fusobacterium nucleatum)参与CRC的发病机制。靶向F. nucleatum的噬菌体也进入了临床试验,利用这些病毒治疗CRC并降低癌症负担。然而,这种单一的靶向也对利用噬菌体治疗没有特异性和有效噬菌体的新兴菌株提出了挑战,这些噬菌体可以通过从野生噬菌体中分离或通过工程设计生成多靶向通用噬菌体来开发。Gogokhia等人报道了从UC患者中分离的有尾病毒目噬菌体可以抑制致癌性粘附侵袭性大肠杆菌的生长,并抑制小鼠模型肠道肿瘤的生长。此外,噬菌体编码一种解聚酶,使它们能够降解生物膜并接近驻留的生物体。也可以设计具有额外治疗优势的噬菌体,例如,使用叠氮化物修饰的噬菌体连接伊立替康-葡聚糖纳米颗粒治疗CT26结直肠癌小鼠,可降低梭杆菌(Fusobacterium spp.)的水平,并有效抑制肿瘤生长。
以上研究强调,需要指出噬菌体疗法在结肠炎和CRC中的潜在治疗作用。噬菌体通常对细菌的靶标范围很窄的主要限制可以通过设计多价、广泛靶标的人工噬菌体来解决,或者储存更多的野生噬菌体用于临床应用。此外,扩大噬菌体疗法在人类治疗IBD和CRC中的应用还需要更大规模的随机对照临床试验。
肿瘤病毒治疗是以溶瘤病毒(Oncolytic viru, OVs)为基础的免疫治疗,通过调节肿瘤微环境(tumor microenvironments, TME)来逆转免疫抑制状态,进而刺激抗肿瘤免疫。OVs是一种天然的或经过基因改造的病毒,旨在靶向并杀死癌细胞,而不会对正常细胞造成明显损害。目前,包括单纯疱疹病毒(herpes simplex virus, HSV)、牛痘病毒(vaccinia virus, VAC)、腺病毒(adenovirus, AdV)、呼肠孤病毒(reovirus, RV)、麻疹病毒(measles virus, MeV)等在内的多种DNA和RNA病毒正迅速成为治疗癌症的新途径。OVs目前通过基因修饰或与其他策略结合来优化,在不伤害健康细胞的情况下提供更大的特异性和对肿瘤的疗效。表2总结了正在进行的CRC患者临床试验。
OVs可以作为癌症疫苗,增加肿瘤特异性T细胞反应。OVs还可以装备免疫刺激分子(如粒细胞-巨噬细胞集落刺激因子,GM-CSF),以提高其免疫激活特性。携带GM-CSF的OVs可促进树突状细胞(DC)的迁移和成熟,最终导致T细胞反应的启动增强,如talimogene laherparepvec(T-VEC)和pexastimgene devacirepvec(Pexa-Vec或JX-594)。Talimogene laherparepvec (T-VEC)系统(HSV)是一种众所周知的治疗修饰病毒。T-VEC已经被美国食品药品监督管理局(FDA)和欧洲药品管理局批准用于治疗转移性黑色素瘤,T-VEC目前正在CRC临床试验中。Pexa-Vec是一种经过修饰的VAC,可编码GM-CSF和β-半乳糖苷酶,以灭活病毒胸苷激酶基因,目前正在进行多项临床试验。
IL-12是Th1型癌症免疫反应的主要调控因子,是另一种用于装备OVs的细胞因子,如TBio-6517(I/II期)和ONCR-177(I期)。LOAd703 (Phase I/II)是一种双臂ADV,由两种肿瘤坏死因子受体(TNF)家族配体CD40L和4-1BBL组成,可刺激T细胞扩张、效应功能获得、存活和T细胞记忆的发展。
Enadenotucirev (EnAd; ColoAd1)是一种复杂的嵌合病毒,由不同的腺病毒血清型重组而成,目前正在进行临床研究。未修饰的病毒,如呼肠孤病毒Pelareorep (Reolysin),也正在用于CRC治疗的临床试验。
此外,OVs可以与免疫检查点抑制剂(immune checkpoint inhibitors, ICIs)或细胞毒性药物结合,通过协同机制实现最有效的癌症免疫治疗,如Pexa-Vec与Tremelimumab(anti-CTLA-4)和Durvalumab(against PD-L1),TBio-6517与Pembrolizumab(anti-PD-1),ColoAd1与nivolumab(anti-PD-1),T-VEC与Atezolizumab(anti-PD-L1),OH2与HX008(anti-PD-L1),ONCR-177与Pembrolizumab。通过这些多管齐下的研究和临床试验,我们预计一些OVs可能能够获得FDA的批准,成功用于癌症治疗的临床应用。
表2 使用溶瘤病毒治疗CRC患者的临床试验。
注:5-FC, 5-氟胞嘧啶; 5-FU, 5-氟尿嘧啶; AdV, 腺病毒; CTLA-4,细胞毒性T淋巴细胞相关抗原4; dsDNA,双链DNA; dsRNA,双链RNA; GM-CSF,粒细胞-巨噬细胞集落刺激因子; HSV,单纯疱疹病毒; i.a.,肝内动脉; i.t., 瘤内; i.v., 静脉注射; MeV, 麻疹病毒; MSS-CRC,微卫星稳定型结直肠癌; PD-1,程序性细胞死亡蛋白1; PD-L1,程序性死亡配体-1; Ref, 参照; RV, 呼肠孤病毒; ssRNA, 单链RNA; TNBC, 三阴性乳腺癌; VAC, 牛痘病毒; VEGF, 血管内皮生长因子; T-VEC, talimogene laherparepvec。
1Clinicaltrials.gov标识符。2黑色素瘤、实体瘤、头颈部鳞状细胞癌、乳腺癌、晚期实体瘤、TNBC、CRC、非黑色素瘤皮肤癌、肝转移瘤。来源: clinicaltrials.gov; 2021年5月评估。
高通量宏基因组测序技术的出现使我们能够了解人类肠道噬菌体和各种病毒群体的复杂性和丰富性。然而,与肠道微生物组的细菌组成部分相比,肠道病毒群几乎被忽视了,这主要是由于可用的病毒鉴定和分类工具有限;也可能是由于对细菌微生物组本身的重大兴趣,所以病毒群的重要性在某种程度上被忽略了。此外,病毒以几种不同的遗传形式存在,它们的主要核酸(RNA或DNA)、链的特征(阳性或阴性)和链的数量(单链或双链)不同。病毒的复杂性对库的准备和测序策略提出了挑战。目前估计只有1%的病毒群被测序,无法归类的序列(分类或功能上)在60% - 90%之间(称为病毒暗物质),有待鉴定。
VLPs组分可以使用透射电子显微镜(TEM)、宏基因组测序或高通量短读长技术(Roche 454、Illumina平台和Ion Torrent平台)进行检测。最近,两种长读长测序技术(Pacific Biosciences和Oxford Nanopore)被开发出来。这些技术可以帮助构建新的大型病毒基因组,获得甲基化模式的信息,并在单个病毒粒子水平上研究种群结构。病毒宏基因组分析工作流程包括质量控制、reads的过滤和修剪、将reads组装成contigs、去除细菌污染、在病毒数据库中对病毒基因组进行比对以及下游分析。已经专门设计了一些软件和数据库来处理高通量病毒群测序数据(表3),这些工具应该在仔细考虑样本类型和科学问题后使用。因此,一个关键的限制是是否有专业的生物信息学家和适当的硬件来执行如此庞大和耗时的分析。
除了病毒宏基因组测序外,未来的互补方法可能包括肠道病毒宏转录组学(RNA-seq)和病毒宏蛋白质组学。上述技术和其他未来发展可能会很好地塑造病毒群研究,以探索肠道病毒群的发病机制及其对人类的潜在有益影响,为设计有效的基于病毒群的肠道和结肠疾病治疗方案提供见解。
表3 病毒宏基因组分析的选用方法和数据库。
尽管取得了这些相对迅速的进展,但我们对肠道病毒特性的理解仍然存在重大差距。(1)虽然病毒参与结肠炎和CRC的发展已经越来越明显,但目前的理论和观察结果需要进一步的检验和验证。(2)随着许多新技术的出现,如核酸测序、组学分析和生物信息学管道,肠道病毒群的特性及其在病理条件发展中的作用仍处于早期阶段。(3)病毒数据库相对较小且不完整(与细菌基因组数据库相比),限制了我们详细分析健康和疾病中的粘膜病毒的能力。(4)有关人类健康的病毒学研究面临着相当大的个体间差异,这些差异可能受到许多因素的影响,如年龄、性别、种族、地理、饮食以及样本收集、储存和处理。
总之,在病毒群研究、认识、疾病相关性和临床应用/治疗方面有许多重要进展,同样也还有许多障碍。在噬菌体治疗和OVs获得广泛临床应用批准之前,必须适当解决上述障碍。尽管存在这些担忧,但病毒疗法可能值得探索,并且通过分子工程可能具有巨大潜力。
本文除了提供一个基本理念,并总结了病毒群与IBD和CRC的联系外,还讨论了噬菌体是一个很有前景的治疗工具,以对抗人类炎症性肠病和结直肠癌的致病菌。噬菌体治疗的随机、安慰剂对照试验已经在IBD和CRC中进行。此外,迫切需要在病毒分离、宏基因组学、富集培养和生物信息学工具方面设计和发明新的方法,以提高我们未来定义和描述病毒的能力,需要进一步的大规模纵向和长期随访前瞻性研究,以及深入的实验调查和验证。该领域的关键方向是确定肠道微生物群与先天性免疫和适应性免疫之间的动态关系(如因果关系)。此外,噬菌体治疗可与细菌或其他微生物组分(通过FMT、益生素和益生菌)一起应用,以改变毒性和免疫原性,有效地驯服人类加剧的炎症疾病,改善人类健康。
附:缩写
5-FC, 5-氟胞嘧啶; 5-FU, 5-氟尿嘧啶; AdV, 腺病毒; AIEC, 粘附侵袭性大肠杆菌; CAC, 结肠炎相关结直肠癌; CD, 克罗恩病; cDNA, 互补DNA; CMV, 巨细胞病毒; CRC, 结直肠癌; CTLA-4,细胞毒性T淋巴细胞相关抗原4; dsDNA,双链DNA; dsRNA,双链RNA; EnAd, Enadenotucirev;FAD,食品药品监督管理局; gDNA, 基因组DNA; GM-CSF,粒细胞-巨噬细胞集落刺激因子; HBV, 乙型肝炎病毒; HCV, 丙型肝炎病毒; HPV, 人乳头瘤病毒; HSV,单纯疱疹病毒; i.a.,肝内动脉; i.t., 瘤内; i.v., 静脉注射; IARC, 国际癌症研究机构; IBD, 炎症性肠病; ICI, 免疫检查点抑制剂; IEC, 肠上皮细胞; JCV, John Cunningham病毒; LPS, 脂多糖; MeV, 麻疹病毒; MMTV, 小鼠乳腺肿瘤病毒; MNoV, 小鼠诺如病毒; MSS-CRC,微卫星稳定型结直肠癌; OV, 溶瘤病毒; PD-1, 程序性细胞死亡蛋白1; PD-L1, 程序性死亡配体-1; Pexa-Vec, pexastimogene devacirepvec; PV,脊髓灰质炎病毒; Ref, 参照; RV, 呼肠孤病毒; ssRNA, 单链RNA; VAC, 牛痘病毒; VEGF, 血管内皮生长因子; T-VEC, talimogene laherparepvec; TEM, 透射电子显微镜; TME, 肿瘤微环境; TNBC, 三阴性乳腺癌; UC, 溃疡性结肠炎; VAC, 牛痘病毒; VEGF, 血管内皮生长因子; VLP, 病毒样颗粒。
不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消
©2012-2023 北京华媒康讯信息技术股份有限公司 All Rights Reserved. 注册地址:北京 联系电话:010-82736610
广播电视节目制作经营许可证 —(京)字第 17437号 京海食药监械经营备20200522号
京ICP备12011723号 京ICP证150092号
 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)






您已认证成功,可享专属会员优惠,买1年送3个月!
开通会员,资料、课程、直播、报告等海量内容免费看!

打赏金额
认可我就打赏我~
1元 5元 10元 20元 50元 其它
打赏作者
认可我就打赏我~
扫描二维码
立即打赏给Ta吧!
温馨提示:仅支持微信支付!
已收到您的咨询诉求 我们会尽快联系您







































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




