编译:微科盟小宇,编辑:微科盟汤貝、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
猪脂肪的过度积累严重影响了现代养猪业的经济效益。在人类和小鼠的研究中都验证了肠道微生物组在宿主能量代谢中的作用。商业杜洛克(Duroc)猪通常采用高能量和高蛋白质的配方饲粮喂养,这种饮食下的肠道微生物群是否以及如何调节猪的脂肪积累在很大程度上是未知的。本研究系统地研究了698头商业杜洛克猪的肠道微生物组与猪瘦肉率(LMP)的相关性,发现Prevotella copri与猪的脂肪积累显著相关。肥猪肠道中的P. copri含量显著较高。高丰度的P. copri与肥胖相关的血清代谢物浓度增加相关,例如,脂多糖、支链氨基酸、芳香族氨基酸和花生四烯酸的代谢产物。宿主肠道屏障通透性和慢性炎症反应也增加。使用无菌小鼠进行的灌胃实验证实,从实验猪中分离的P. copri是一种增加宿主脂肪积累和改变血清代谢物的致病物种。P. copri灌胃小鼠的结肠、脂肪组织和肌肉转录组数据表明,P. copri定殖通过TLR4和mTOR信号通路激活宿主慢性炎症反应,显著上调与脂肪生成和脂肪积累相关的基因表达,但降低与脂肪分解、脂质转运和肌肉生长相关的基因表达。综上所述,研究结果表明,商业配方饲料喂养的猪肠道微生物群落中的P. copri通过TLR4和mTOR信号通路激活代谢产物引起的宿主慢性炎症反应,并显著增加宿主脂肪积累。该结果为通过调节肠道微生物组成来减少猪脂肪积累提供了基础知识。
原名:Prevotella copri increases fat accumulation in pigs fed with formula diets
译名:Prevotella copri 增加饲喂配方饲粮的猪的脂肪积累
期刊:Microbiome
IF:14.650
发表时间:2021.8.21
通讯作者:黄路生&陈从英
通讯作者单位:江西农业大学猪遗传改良与养殖技术国家重点实验室

过量脂肪积累显著降低了猪瘦肉率(LMP)。因此,在本研究中,我们使用LMP作为一个指标来评估肠道微生物组在猪肥胖中的作用。我们记录了两个农场饲养的698头商业杜洛克猪的LMP。发现群组包括来自两个农场(280头来自沙湖,270头来自江阴)的550头猪(309头雄性和241头雌性),验证群组包括来自江阴牧场的148头猪(100只雄性和48只雌性)。表型值基本符合正态分布(图S2)。所有的698头猪在160天龄时收集了粪便样本,我们对其16S rRNA基因进行了高变区测序(发现群组为V4区,验证群组为V3–V4区)。我们首先分析了发现群组中OTUs的肠型和共丰度组(CAGs)与LMP的关系。所有样本均聚集成两个以Prevotella或者Treponema为优势菌属的肠型组,且Prevotella肠型猪的LMP显著降低。(图S3)。在CAG水平,共有1159 OTUs被用于构建共丰度网络。根据SparCC相关系数将这些OTUs分为12个共丰度组(CAGs)。(图1a,图S4)。CAG3含有的OTUs主要注释为Prevotella,尤其是P. copri,与LMP呈负相关;而CAG8含有的OTUs主要注释为F. prausnitzii和R. flavefaciens,与LMP呈显著正相关,表明这些CAGs在与LMP相关的肠道菌群功能行会中的核心作用(图1a)。我们采用两部分模型进行了全细菌相关性研究,以确定发现群组中与LMP显著相关的细菌类群。FDR < 0.01时共鉴定出166个与LMP相关
OTUs
,其中82个OTUs与LMP呈正相关,84个OTUs与LMP呈负相关。正相关的OTUs大多属于梭菌目(Clostridiales),如F. prausnitzii、Lachnospiraceae和Ruminococcaceae,但负相关的OTUs以
普雷沃氏菌
(Prevotella)为主(40/84)。其中18个P.copriOTUs与LMP呈最强的负相关(图1b,表S2)。
肠型分析的结果在验证群组中得到了很好的重复。属于普雷沃氏菌的
OTUs
(如P. copri)聚集在与LMP负相关的CAG8,注释到P. copri的OTU17是共丰度网络中的枢纽节点(图S5)。在验证群组中共识别出11个
LMP
相关OTUs。大多数注释到的LMP相关OTUs的细菌类群与发现群组中的相同(8/11)。与LMP正相关的5个OTUs中的2个被认为是属于Christensenellaceae。而与LMP呈最显著负相关的OTUs被注释为Prevotella和
P. copri
(表S3)。这些结果进一步表明,P. copri与猪的脂肪积累显著相关。我们进一步对验证群组中包含的16个粪便样本进行鸟枪宏基因组测序,包括8个LMP值最高的样本和8个LMP值最低的样本(图S6a)。与之前在人类身上的发现一致,肥猪肠道微生物组中的基因数量和α多样性(Shannon指数)明显低于瘦猪(图S6b,c).我们通过LEfSe分析确定了40个菌种与LMP相关。在肥猪中富集的细菌种类中普雷沃氏菌占主导地位(13/20)。尤其是P. copri与LMP呈最强的负相关,是降低LMP的细菌物种中的中心物种(图S6d)。总共有20个菌种在瘦猪中富集,其中大部分是来自Treponema和Clostridiales的产生SCFAs的细菌,例如Lactobacillus reuteri和Bifidobacterium longum(图1c)。为了扩展宏基因组测序数据,我们整合了20份来自发现群组粪便样本的宏基因组测序数据,这些样本是我们之前研究中根据饲料效率的表型值选择的。与16个样本的结果相似,综合36个样本的结果表明,包括P. copri在内的4种普雷沃氏菌与LMP降低显著相关,而Treponema和Clostridiales在瘦猪中的丰度较高(图S7a)。
图1 P. copri是一种降低猪瘦肉率(LMP)的枢纽细菌。a OTU水平网络图。节点大小显示每个OTU的平均数量。节点之间的红线和灰线分别表示节点之间的正相关性和负相关性。仅显示相关性大于0.5的分类单元。当P < 0.005时,经过质量控制的OTUs通过置换多变量方差分析(PERMANOVA)分组为12个CAGs。右侧的颜色渐变显示CAGs和LMP间相关性的P值和系数。b与猪LMP相关的粪便微生物群的关键OTUs。c利用宏基因组测序数据发现与LMP显著相关的细菌物种。接下来,我们通过使用宏基因组测序数据将微生物基因目录映射到CAZy和KEGG模块,研究了与LMP相关的肠道微生物组的功能能力。我们共鉴定出50种CAZymes,它们在肥猪和瘦猪之间具有显著不同的丰度,其中27种CAZymes涉及在瘦猪中富集的半乳糖、木聚糖和甘露糖的代谢。肥胖个体肠道微生物群中丰度显著较高的23种CAZymes(如,GH28、PL11、GH22、PL10和GT4)主要参与鼠李糖和葡聚糖的代谢以及脂多糖的生物合成(图2a)。LMP相关的OTUs和CAZ
ymes
之间的相关分析表明,LMP相关细菌对CAZymes的变化有贡献(图2b)。与LMP相关的KEGG路径如图2c。在肥猪的肠道微生物群中,包括脂多糖生物合成和花生四烯酸代谢,参与介导炎症反应的17个KEGG通路的丰度显著较高;FoxO信号通路、胰岛素抵抗、BCAA(缬氨酸、亮氨酸和异亮氨酸)生物合成以及与肥胖和胰岛素抵抗相关的芳香族氨基酸(酪氨酸和苯丙氨酸,AAA)代谢,以及双组分系统、细菌趋化性、鞭毛装配,碳水化合物的消化和吸收与从细菌获取能量的能力增加有关。与瘦猪相比,肥猪体内细菌侵入上皮细胞的通路(该途径会损害肠道屏障的完整性)也高度富集(图2c)。所有这些通路都与肥胖相关的细菌种类,尤其是
P. copri
呈正相关(图2d),表明肥猪的细菌种类可能会产生更多与炎症反应、肥胖和胰岛素抵抗相关的因子,损害宿主肠道屏障完整性,并提高能源收集能力。我们进一步从低LMP值的实验猪的粪便样本中
分离和培养
P. copri
。使用
Nanopore
平台进行全基因组测序。P. copri的基因组全长3.44 Mb,包含3039个编码基因(CDS)(图S8)。我们首先根据P. copri分离株的基因组序列构建了系统发育树,包括来自西化人群的60个分离株,非西化人群的51个分离株,以及本研究中的1个猪分离株。在本研究中,从猪中分离到的P. copri菌株明显位于西化中国人群的分支中(图3a)。共鉴定出24个多糖利用位点(PULs),超过10个PULs在西化人群中的患病率较高(图S9)。在该P. copri分离株的基因组中发现了涉及花生四烯酸代谢、BCAA生物合成、AAA生物合成和代谢、TCA循环和蛋白质输出的基因,并结合宏基因组测序数据确定了这些基因在受试样品中的丰度。与仅通过宏基因组测序数据确定的LMP相关功能能力相一致,肥猪的肠道微生物群具有显著较高的参与花生四烯酸代谢、BCAA生物合成、AAA生物合成和代谢以及胰岛素抵抗的P. copri基因丰度,但与瘦猪相比,参与TCA循环和蛋白质输出的基因丰度较低(图3b)。考虑到肥猪P. copri的高丰度,P. copri是与LMP相关的宏基因组功能能力变化的主要贡献者。相反地,15个KEGG通路在瘦猪中的丰度显著较高(图2c),包括丁酸代谢、柠檬酸循环(TCA循环)、辅助因子和维生素代谢、赖氨酸降解、半胱氨酸和蛋氨酸代谢、精氨酸和脯氨酸代谢。所有这些通路与多个高LMP相关细菌物种呈正相关(图2d)。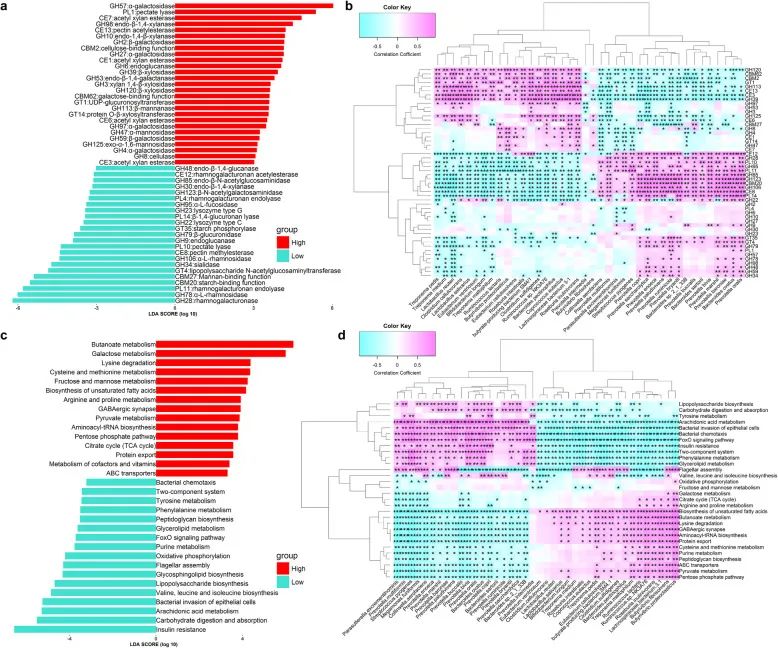 图2 利用宏基因组数据研究与猪瘦肉率(LMP)相关的肠道微生物群的功能能力及其与LMP相关细菌的相关性。a 与LMP相关的CAZy功能模块。b LMP相关CAZymes和LMP相关细菌种类的相关性。c 与LMP相关的KEGG通路的功能模块。d LMP相关KEGG通路和LMP相关细菌种类的相关性。LMP相关功能模块与细菌种类之间的相关性设定在阈值| r | > 0.5,FDR < 0.05
图2 利用宏基因组数据研究与猪瘦肉率(LMP)相关的肠道微生物群的功能能力及其与LMP相关细菌的相关性。a 与LMP相关的CAZy功能模块。b LMP相关CAZymes和LMP相关细菌种类的相关性。c 与LMP相关的KEGG通路的功能模块。d LMP相关KEGG通路和LMP相关细菌种类的相关性。LMP相关功能模块与细菌种类之间的相关性设定在阈值| r | > 0.5,FDR < 0.05
 图3 本研究中分离到的P. copri基因组信息。a 西化和非西化人群P. copri分离株以及本研究中基于基因组序列的P. copri的系统发育树。外圆的不同颜色代表样品来自的国家;内圈的不同颜色表示饮食。b 通过整合宏基因组测序数据,比较P. copri基因组中确定的基因丰度,这些基因涉及花生四烯酸代谢、BCAA生物合成、AAA生物合成和代谢、胰岛素抵抗、柠檬酸循环以及瘦肉和肥肉之间的蛋白质输出。高:肥猪;低:瘦猪 * FDR < 0.05,** FDR< 0.01,*** FDR < 0.005。
图3 本研究中分离到的P. copri基因组信息。a 西化和非西化人群P. copri分离株以及本研究中基于基因组序列的P. copri的系统发育树。外圆的不同颜色代表样品来自的国家;内圈的不同颜色表示饮食。b 通过整合宏基因组测序数据,比较P. copri基因组中确定的基因丰度,这些基因涉及花生四烯酸代谢、BCAA生物合成、AAA生物合成和代谢、胰岛素抵抗、柠檬酸循环以及瘦肉和肥肉之间的蛋白质输出。高:肥猪;低:瘦猪 * FDR < 0.05,** FDR< 0.01,*** FDR < 0.005。
3 肥猪血清代谢组的变化及其与肠道菌群变化的相关性我们首先使用酶联免疫吸附试验(ELISA)测定并比较了瘦猪(n=8)和肥猪(n=8)血清LPS和脂多糖结合蛋白(LBP)的浓度。与肠道微生物群中脂多糖生物合成功能的丰富性相一致,肥猪血清脂多糖浓度显著高于瘦肉猪(P < 0.005)。LBP也观察到同样的结果(P <0.05;图4a)。随后,我们确定了从验证群组中随机收集的38份血清样本的非靶向代谢组谱。通过Spearman秩相关分析(FDR<0.05),我们确定了80个代谢产物特征与LMP显著相关(表S5)。这些LMP相关代谢物聚集在23条KEGG通路中,涵盖大部分与LMP相关的肠道微生物功能通路(图S10)。接下来,我们重点关注一些基于肠道微生物组LMP相关功能能力的有趣的LMP相关代谢物。肥猪的血清BCAA、AAA及其相关代谢物浓度明显高于瘦猪(图4b和表S5)。与瘦猪相比,肥猪的血清中与炎症反应和代谢综合征相关的代谢物浓度显著升高,如
3-
甲基-2-氧代戊酸,一种BCAA代谢的中间产物,可诱导BCAA,L-鼠李糖(脂多糖的重要成分)和花生四烯酸代谢的代谢物(5-HETE、9-HETE、白三烯和前列腺素)的积累(图4b,表S5)。这与预测的肥猪肠道微生物组对BCAA生物合成、AAAs和花生四烯酸代谢的功能能力显著一致(图2b)。和肥猪相比,瘦猪具有更高浓度的代谢物,这些代谢物已报道可以减少猪的脂肪积累并增加血清中的瘦肉肌肉质量。例如,肌酸、甜菜碱代谢产物(L-组氨酸,三甲基甜菜碱和脯氨酸甜菜碱)和抗炎因子(脂蛋白A4)(图4b,表S5)。
儿茶酚胺,包括多巴胺、
N-乙酰多巴胺和L-dopa在瘦猪血清中也表现出较高的丰度,儿茶酚胺通过增加脂肪分解来减少脂肪组织中的脂肪积累,从而减少脂肪生成并促进游离脂肪酸(FFA)的运输,(表S5和图4b)。在38个有代谢组数据的样本中,我们进一步评估了LMP相关细菌在OTU水平上对血清代谢物变化的贡献
(
图S11)。在种水平上,16个样本同时具有宏基因组测序和代谢组学数据(图4c)。P. copri与上述几乎所有与肥胖相关的代谢产物呈正相关,与瘦肉相关的代谢产物呈负相关。其他LMP相关细菌也在不同程度上对LMP相关代谢物有贡献。例如,P. copri和两个拟杆菌与血清BCAA浓度均显著相关,但BCAA更大程度上是由P. copri驱动的(图S12)。这些结果表明,LMP相关细菌对与脂肪积累相关的宿主血清代谢物的转移有重要作用。综上所述,慢性炎症相关代谢物,例如,BCAA、AAA和花生四烯酸的代谢物在肥猪体内含量较高,而与抗炎、脂质代谢和能量消耗相关的代谢物在瘦猪血清中含量较高。在肥猪中,P. copri和其他LMP相关细菌对血清代谢物的变化作出响应。 图4 脂多糖(LPS)、脂多糖结合蛋白(LBP)和部分血清代谢物在肥瘦猪之间的标准化丰度差异,以及它们与LMP相关细菌种类的相关性。a 肥猪和瘦猪血清LPS和LBP浓度的比较。b 高LMP(n=8)和低LMP(n=8)猪之间标准化丰度差异的代谢物。红色条表示瘦猪代谢产物的标准化丰度,蓝色条表示肥猪代谢产物的标准化丰度。c 肥胖相关代谢物与LMP相关细菌种类之间的相关性。d 高LMP相关代谢物与LMP相关细菌种类之间的相关性。颜色条表示相关系数的值。
图4 脂多糖(LPS)、脂多糖结合蛋白(LBP)和部分血清代谢物在肥瘦猪之间的标准化丰度差异,以及它们与LMP相关细菌种类的相关性。a 肥猪和瘦猪血清LPS和LBP浓度的比较。b 高LMP(n=8)和低LMP(n=8)猪之间标准化丰度差异的代谢物。红色条表示瘦猪代谢产物的标准化丰度,蓝色条表示肥猪代谢产物的标准化丰度。c 肥胖相关代谢物与LMP相关细菌种类之间的相关性。d 高LMP相关代谢物与LMP相关细菌种类之间的相关性。颜色条表示相关系数的值。
鉴于肥猪血清LPS和与炎症反应相关的代谢物浓度增加,为了检测宿主肠道屏障通透性和慢性炎症反应,我们测定了血清中生物标志物连蛋白,二胺氧化酶(DAO),FABP2,促炎细胞因子肿瘤坏死因子α(TNFα),白细胞介素-1β(IL-1β),白细胞介素-6(IL-6)和干扰素-γ(IFN-γ)。如预期的那样,与高瘦猪(n=8)相比,肥猪(n=8)血清中的DAO未发现显著差异,但连蛋白(P < 0.005)和FABP2(P=0.083)浓度更高(图5a),表明肥猪的肠道屏障通透性增加。此外,肥猪的血清TNF-α、IL-1β、IL-6和IFN-γ浓度也较高(图5b),表明宿主有慢性炎症反应。综上所述,代谢组学分析结果表明,肠道高丰度P. copri菌可能通过LPS、BCAA、AAA和花生四烯酸等代谢产物诱导宿主肠道屏障通透性并促进慢性炎症反应,从而增加猪的脂肪积累。
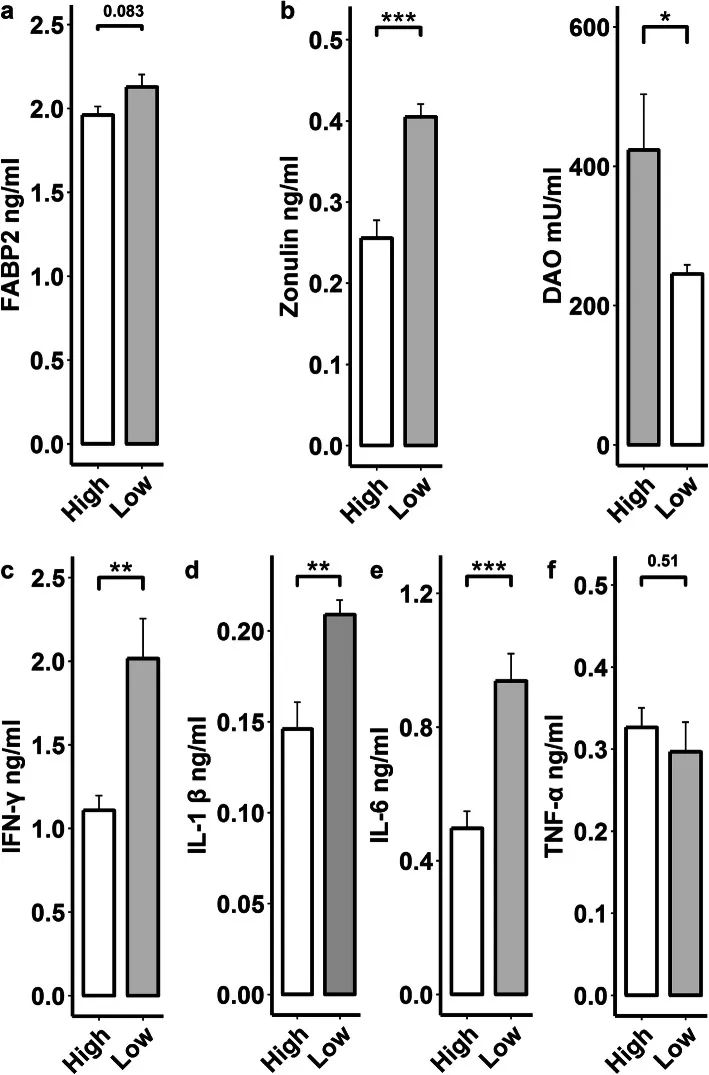
图5 比较高(n=8)和低(n=8)LMP猪的肠道通透性病理生理上皮完整性血液标志物(脂肪酸结合蛋白2(FABP2)、连蛋白和二胺氧化酶(DAO))以及促炎细胞因子的血清浓度。a-b 瘦猪和肥猪之间FABP2、连蛋白和DAO血清浓度的比较。c-f 瘦猪和肥猪血清促炎细胞因子(IL-1β、IL-6、TNF-α和IFN-γ)浓度的比较。* P < 0.05,** P < 0.01,和*** P < 0.005。
接下来,我们通过在无菌小鼠中使用P. copri菌进行灌胃实验,来评估从实验猪中分离的P. copri菌与宿主脂肪积累之间的可能因果关系。在1个月灌胃实验结束时对粪便DNA进行的qPCR分析证实,在每只接受治疗的小鼠的肠道中成功地定殖了P. copri(图6a)。表型测定发现,饲喂正常饲料的P. copri灌胃小鼠的体脂率(P< 0.01)和附睾脂率(P < 0.05)显著增加(图6b)。在喂食高脂饮食(HFD)的P. copri灌胃小鼠中,脂肪积累变得更加严重(P < 0.005)(图6b)。
灌胃细菌显著提高了血清LPS和LBP浓度(P < 0.005)。此外,HFD可提高LPS的血清浓度(图6c)。P. copri定殖也导致血清中肠屏障通透性生物标记物FABP2(P < 0.01)和连蛋白(P < 0.05)浓度升高。值得注意的是,饲喂高脂肪食物的小鼠增强了肠道屏障的通透性。在P. copri定殖并喂食高脂肪饮食的小鼠中,DAO的血清浓度增加近两倍(图6d)。P. copri定殖小鼠的促炎细胞因子(IL-6、IL-1β、TNF-α、和IFN-γ)的血清浓度显著高于PBS灌胃对照小鼠。通过喂养高脂肪食物给P. copri定殖小鼠,这种情况得到了加强(P < 0.01))(图6e)。
我们共鉴定出222种血清代谢物,在对照组和P. copri定殖小鼠之间的丰度存在显著差异,以及对照组和喂食HFD的P. copri定殖小鼠之间215种差异代谢物特征(表S6)。对照组和P. copri定殖小鼠之间的差异代谢物富集到与瘦猪和肥猪之间高度相似的通路(图S13)。例如,P. copri定殖小鼠(在正常食物和HFD中)增加了与BCAA生物合成、AAA代谢和花生四烯酸代谢相关的通路的丰富度。生物素代谢通路、丁酸代谢通路和泛酸通路在P. copri定殖小鼠中的丰度较低(图6f)。而且,正如在猪上观察到的,参与炎症反应和代谢综合征的代谢物,如BCAA、AAA、白三烯、前列腺素、HETE和L-鼠李糖在P. copri定殖小鼠中含量较高,而代谢物与瘦肌肉质量、能量消耗和减少脂质积累有关(例如甜菜碱、维生素和多巴胺)在P. copri定殖小鼠的血清中较低(图6g和表S6)。
 图6 确认P. copri在宿主脂肪积累中的因果作用。a qPCR证实了P. copri在无菌小鼠中的成功定殖。Y轴表示RQ值,该值反映了灌胃小鼠中P. copri的相对丰度。b比较P. copri灌胃小鼠(n=7)、喂食高脂饮食的P. copri灌胃小鼠(n=7)和对照小鼠(n=7)的体脂百分比(%)和附睾脂肪百分比(%)。c 比较实验小鼠组中LPS内毒素和LBP的血清浓度。d 实验小鼠组间FABP2、zonulin和DAO血清浓度的比较。e 比较实验小鼠组间促炎细胞因子(IL-1β、IL-6、TNF-α、ANDIFN-γ)的血清浓度。f 实验小鼠组间差异血清代谢物富集的KEGG通路的比较。红色圆圈代表实验小鼠组,红色圆圈的大小表示差异代谢物的富集。实验小鼠组中标准化丰度存在差异的代谢物,* P < 0.05,** P < 0.01,和*** P < 0.005。
图6 确认P. copri在宿主脂肪积累中的因果作用。a qPCR证实了P. copri在无菌小鼠中的成功定殖。Y轴表示RQ值,该值反映了灌胃小鼠中P. copri的相对丰度。b比较P. copri灌胃小鼠(n=7)、喂食高脂饮食的P. copri灌胃小鼠(n=7)和对照小鼠(n=7)的体脂百分比(%)和附睾脂肪百分比(%)。c 比较实验小鼠组中LPS内毒素和LBP的血清浓度。d 实验小鼠组间FABP2、zonulin和DAO血清浓度的比较。e 比较实验小鼠组间促炎细胞因子(IL-1β、IL-6、TNF-α、ANDIFN-γ)的血清浓度。f 实验小鼠组间差异血清代谢物富集的KEGG通路的比较。红色圆圈代表实验小鼠组,红色圆圈的大小表示差异代谢物的富集。实验小鼠组中标准化丰度存在差异的代谢物,* P < 0.05,** P < 0.01,和*** P < 0.005。
6 对P. copri灌胃小鼠结肠、脂肪和肌肉组织的转录组分析阐明了肠道微生物组影响宿主脂肪积累的机制为了确定P. copri影响宿主体脂肪百分比的分子机制,对从喂养正常食物的对照组和定殖小鼠身上获取的结肠、白色脂肪和肌肉组织进行了RNA测序分析。在对照组和大肠杆菌定殖小鼠的结肠、白色脂肪和肌肉组织中分别鉴定出225、338和384个差异表达基因(DEGs) (FDR < 0.05)(表S7)。根据血清LPS、LBP、BCAA和促炎细胞因子浓度的增加,我们特别关注分别对细菌LPS和BCAA产生响应的TLR4和mTOR信号通路中的基因表达。正如所料,与对照组相比,在结肠和附睾脂肪组织中,P. copri定殖小鼠TLR4、CD14、Myd88、Mal、Irak1、Irak2和Irak4的表达水平较高(FDR < 0.05),但在肌肉中没有这种表达趋势(图7a)。这表明结肠和白色脂肪组织中TLR4信号通路的激活。mTOR信号通路也被激活。Mlst8是mTORC1的核心成分,在结肠组织中上调。Rheb、Pdk1、Atg 13和Atg101在所有三种组织中均表达上调。然而,Tsc2(mTORC1的调节剂)和Deptor(mTORC1的抑制亚单位)在所有三种组织中均下调。先前的研究表明,导致脂质积累的mTORC1的激活与Hif1a的上调密切相关。有趣的是,Hif1a在所有三种组织中均上调(图7b)。
我们还发现了一些与免疫和炎症反应、脂肪积累和肥胖相关的有趣的DEGs。在P. copri定殖小鼠的结肠组织中,参与免疫和炎症反应的基因(例如,Ccl2、Ccl24、Ccl3、Ccl4、Ccl7、Il1b、Il6ra、Ilf2、Tlr2、Tlr3、Tlr5和来自免疫球蛋白超家族的六个基因)和脂肪积累和肥胖相关的基因(如Fabp2和Ins2)的mRNA水平较高(图7c,表S7)。这与P. copri引起的慢性炎症反应和肠道通透性增加相一致。在白色脂肪组织中,参与脂肪生成(Fabp9、Scd1、Scd2、和Scd3)和炎症反应(Il13ra2)的DEG在
P. copri
定殖小鼠中的表达水平高于对照小鼠。然而,一些与脂解和脂质转运相关的基因(Abca1、Apoc1、Apoe、Pparg、Cpt2、和Adrb3)在P. copri定殖小鼠中的表达水平较低(图7c)。在肌肉中,有几个与脂肪生成和脂质沉积有关的基因(如Adipoq、Adipor2、Apobr、Dgat2、Fabp3、Scd2、Pck1、Ppargc1a)和炎性反应(Il11ra2、Il6ra、Ilf2)在P. copri定殖小鼠中表达水平较高,而与骨骼肌生长相关的Igf2r和Igfbp7基因在P. copri定殖小鼠中表达水平较低(图7c)。综上所述,结果表明P. copri定殖激活了TLR4和mTOR信号通路,并上调了与免疫和炎症反应相关的基因以及与脂肪生成和脂肪积累相关的基因的表达水平,同时下调与脂解、脂质转运和肌肉生长相关的基因表达水平。 图7 P. copri诱导宿主结肠、脂肪和肌肉组织基因表达水平的变化。a TLR4信号级联的基因差异表达水平。b mTOR通路中基因表达的变化。c 在对照组和P. copri定殖小鼠之间,与结肠、白色脂肪组织和肌肉中涉及免疫、脂解、脂质转运和肌肉生长相关基因的差异表达水平。*P < 0.05; ** P <0.01,和*** P < 0.005。
图7 P. copri诱导宿主结肠、脂肪和肌肉组织基因表达水平的变化。a TLR4信号级联的基因差异表达水平。b mTOR通路中基因表达的变化。c 在对照组和P. copri定殖小鼠之间,与结肠、白色脂肪组织和肌肉中涉及免疫、脂解、脂质转运和肌肉生长相关基因的差异表达水平。*P < 0.05; ** P <0.01,和*** P < 0.005。
7
饲粮对本研究分离的
P. copri
定殖的影响以往的研究表明,不同的饮食习惯会影响肠道P. copri的基因组功能和菌株表达。我们首先使用来自工业养猪环境下的野猪(n=6;自由生活,高纤维饲料)、藏猪(n=13;半放牧,高纤维饲料)、上述杜洛克猪(n=20和16;高能量和蛋白质配方饲料)和杜洛克×(长白猪×约克郡)猪(DY,n=20)的宏基因组测序数据比较了在不同饲养模式下的猪内脏中的P. copri的丰度。杜洛克猪(2.68%±0.49%(SE)和1.20%±0.28%,总的映射到P. copri的reads与总clean reads的百分比)和DLY(2.67%±0.41%)与野猪(0.15%±0.05%)和藏猪(0.23%±0.03%)相比,观察到显著更高的P. copri丰度(图8a,P<0.05)。
我们进一步研究了饮食对C57BL6无菌小鼠P. copri定殖的影响。18只无菌小鼠分为三组,分别喂食标准食物、高脂肪食物或高纤维食物,并灌胃P. copri。高脂饲料组小鼠的定殖P. copri丰度极显著高于对照组( P < 0.01),而标准饲料组和高纤维饲料组小鼠的定殖P. copri丰度差异不显著(图8b)。此外,与喂食标准饲料的小鼠相比,喂食高脂肪饲料的小鼠体脂和附睾脂肪的百分比均显著升高( P < 0.005)。高纤维饲料喂养的小鼠比标准饲料喂养的小鼠脂肪积累少(P <0.05),但差异不显著(图8c,图S14)。这应该是由于与标准饮食组相比,高纤维饮食组的饮食含有较少的碳水化合物和能量。至于饮食对P. copri定殖小鼠宿主肠道屏障通透性和慢性炎症反应的影响,喂食标准食物和喂食高纤维饮食的P. copri大鼠之间,LPS、LBP、生物标记物连蛋白和FABP2、促炎细胞因子(IL-1β和IL-6)或TNF-α的浓度没有显著差异(图S14)。然而,高脂饮食显著增强了P. copri诱导的宿主肠道屏障通透性和慢性炎症反应(P < 0.05,图S14)。
 图8 饮食对肠道P. copri丰度的影响。a 在工业养猪环境下,比较野猪、藏猪、杜洛克猪和杜洛克×(长白猪×约克郡)猪(DLY,n=20)内脏中的P. copri丰度。**P < 0.01, ***P < 0.005, ****P < 0.001。b 饮食对P. copri灌胃小鼠脂肪积累的影响。将由P. copri定植的C57BL6无菌小鼠随机分为三组,分别喂食标准食物(CD,n=6)、高脂肪饮食(HFD,n=6)和高纤维饮食(纤维,n=6)。
图8 饮食对肠道P. copri丰度的影响。a 在工业养猪环境下,比较野猪、藏猪、杜洛克猪和杜洛克×(长白猪×约克郡)猪(DLY,n=20)内脏中的P. copri丰度。**P < 0.01, ***P < 0.005, ****P < 0.001。b 饮食对P. copri灌胃小鼠脂肪积累的影响。将由P. copri定植的C57BL6无菌小鼠随机分为三组,分别喂食标准食物(CD,n=6)、高脂肪饮食(HFD,n=6)和高纤维饮食(纤维,n=6)。
已有的证据表明,肠道微生物群可能有助于宿主脂肪的积累。在这项研究中,我们从配方饲料喂养的猪的肠道微生物群中鉴定出的P. copri是一种增加猪脂肪积累的中枢细菌。P. copri是由几个不同的分支组成的复合体。根据习惯性饮食,它与宿主健康既有积极的联系,也有消极的联系。例如,在喂食富含纤维的饮食的小鼠中,P. copri的定殖通过肠道糖异生改善了葡萄糖稳态。普氏菌丰度或普氏菌-拟杆菌比例能预测食用全谷物或高纤维饮食的超重参与者的体重和脂肪减肥成功与否。然而,一项临床试验报告显示,随着肠道通透性的增加,循环琥珀酸水平升高,Prevotellaceae和Veillonellaceae的相对丰度较高,这与肥胖和葡萄糖代谢受损有关。2型糖尿病患者粪便中
P. copri
的患病率和血浆白细胞介素6水平升高。P. copri与人类胰岛素抵抗和加重葡萄糖不耐受有关。不同的习惯性饮食导致人类肠道P. copri菌株具有不同的遗传和功能特征,并且人类肠道P. copri菌株显示出不同的多糖利用谱。在现代工业化的养猪中,为了最大限度地开发猪的生长潜力,人们向猪群提供经过加工、含有大量可消化能量和蛋白质的商品配方饲粮。这些日粮选择并塑造了商业猪的肠道P. copri。与工业养猪环境下的杜洛克猪和杂交DLY猪相比,杜洛克猪和杂交DLY猪的饲料中含有高消化能量和高蛋白质含量,喂饲高纤维饲料的野猪和藏猪的P. copri丰度显著降低。此外,通过对无菌小鼠的灌胃实验,我们观察到在喂食高脂肪饮食的小鼠中分离出的P. copri的丰度显著增加。来自发现群组的杜洛克猪的肠道微生物组比来自验证群组的猪的P. copri丰度更高。在一个群组中为猪提供了相同的配方饲料,但我们观察到两个实验群组中的肠道
P. copri
丰度存在显著差异。这可能是由母体效应或/和性能测量前的饮食造成的(从出生到30公斤体重)。两组实验猪均来自不同环境和管理模式的不同养殖场。发现群组配方饲料的消化能量和粗蛋白质高于验证组杜洛克猪(3023对2960 kcal/kg,17%对15%)。此外,这些猪在基因上是不相关的(来自不同的母猪)。宿主遗传学可能是导致肠道内P. copri丰度显著变化的另一个原因。肥猪血清LPS、LBP、BCAA、AAA及花生四烯酸代谢相关代谢产物浓度显著高于瘦猪。先前在小鼠身上的报道也指出了肠道细菌脂多糖在肥胖中的作用。在人类身上的几项研究表明,P. copri在很大程度上促进了微生物对支链氨基酸生物合成潜力的增加,这些研究表明,血清BCAAs水平或其分解产物在2型糖尿病、肥胖和胰岛素抵抗中起着因果作用。花生四烯酸是合成一系列生物活性化合物的底物,包括前列腺素和白三烯。这些化合物可以作为炎症细胞因子产生和免疫功能的介质和调节器。至于AAAs,据报道,AAAs循环浓度的增加与人类肥胖和胰岛素抵抗有关。猪的LMP相关细菌分类群与血清代谢物之间的相关性表明,肠道微生物群,尤其P. copri,导致血清BCAA、AAA和花生四烯酸代谢物水平升高,这意味着肠道微生物群应通过这些代谢物诱导慢性炎症反应,从而导致宿主脂肪积累。相比之下,据报道在人类中具有抗炎作用的细菌种类在瘦猪中显著富集,包括
F. prausnitzii
,在两个实验猪群中,其在肠道中的丰度与
P. copri
呈负相关
(图
S15
)
。在瘦猪的肠道微生物群中,辅助因子和维生素代谢的功能项得到了丰富。瘦猪血清中富含维生素
K
、
D3
、泛酸和生物素,据报道,这些维生素与减少普通肥胖、减少炎症以及增加能量消耗和脂联素表达有关。在我们之前的研究中,我们发现在本研究的发现群组280头杜洛克猪中,Prevotella肠型的平均日采食量高于Treponema肠型。Prevotella(主要是P. copri)可能是与宿主采食量相关的关键细菌种类。总的来说,结合本研究和Yang等人在猪身上的实验结果以及P. copri定殖小鼠的结果,我们提出了肠道微生物组对宿主脂肪积累影响的模型:(1)肠道中大量的Prevotella,尤其是
P. copri
,可能与能量摄入过多有关;(2)P. copri导致血清LPS、BCAA和花生四烯酸代谢物浓度增加,激活TLR4和mTOR信号通路,导致宿主慢性炎症反应;(3)与脂肪生成和脂肪积累相关的基因上调,而与脂肪分解、脂肪转运和肌肉生长相关的基因下调,这将增加脂肪积累并降低LMP。综上所述,我们从喂食商业配方饲料猪的肠道微生物群中鉴定并证实了P. copri显著增加猪的脂肪积累。高丰度的肠道P. copri通过TLR4和mTOR信号通路激活代谢物的宿主慢性炎症反应,并导致脂肪积聚。研究结果为通过调节养猪业中的肠道微生物组成来减少猪的脂肪积累和增加LMP提供了基础知识,并为肠道微生物组群对人类肥胖的影响提供了参考见解。




 京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)
京公网安备 11010802020745号
工商备案公示信息
互联网药品信息服务资格证书((京)-非经营性-2020-0015)







































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612




