编译:微科盟Erics,编辑:微科盟木木夕、江舜尧。
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》
导读
当前对沿海沉积物岩心中反硝化菌和厌氧氨氧化菌的活动,丰度和群落组成与深度变化的关联研究仍然非常有限。本研究结合15N同位素标记诱导实验、定量聚合酶链反应(qPCR)和高通量测序技术探讨了冬季和夏季收集的大亚湾沉积物岩心(大约100 cm深)中反硝化菌和厌氧氨氧化菌的结构和功能。结果表明,即使在较深的沉积物中,溶解无机氮(DIN)的浓度较低的情况下,也能检测到反硝化菌和厌氧氨氧化菌的活性和丰度。反硝化菌和厌氧氨氧化菌的潜在比率,丰度和群落组成仅在空间上有所变化。在表层沉积物中(顶部2 cm),反硝化菌的活性和丰度明显高于厌氧氨氧化菌,但在底部沉积物中,厌氧氨氧化菌对氮损失的相对贡献度增加到60%以上。系统发育分析表明,nirS型反硝化菌属于10个不同的集群,念珠菌在整个沉积物中占据厌氧氨氧化菌群的主导地位。此外,表下沉积物中的反硝化菌和厌氧氨氧化菌均不同于表层沉积物。硝化和反硝化或厌氧氨氧化耦合可能在去除固定氮中起重要作用,电子受体(例如亚硝酸盐和硝酸盐)的可用性强烈影响了底部沉积物中氮的损失,强调了其作为埋藏氮汇的作用。
原名:Variations in nitrogen removal rates and microbial communities over sediment depth in Daya Bay, China
译名:大亚湾沉积物深度对氮去除速率和微生物群落的影响
期刊:Environmental Pollution
IF:8.071
发表时间:2021.04.30
通讯作者:洪义国
通讯作者单位:广州大学环境科学与工程学院
本研究使用重力取芯器从4个代表性地点(S1、S5、S8和S13)收集中国南海大亚湾(113°29ʹ42′′–114°49ʹ42′′E和23°31ʹ12′′–24°50ʹ00′′N)沉积物岩心。该海湾是中国南海最大和最重要的半封闭海湾之一,南北长约24公里,宽13-25公里,面积约600平方公里。采样时间为2015年8月(夏季)和12月(冬季)。结合15N同位素标记诱导实验、定量聚合酶链反应(qPCR)和高通量测序技术揭示冬季和夏季收集的大亚湾沉积物岩心(约100 cm深)中反硝化菌和厌氧氨氧化菌的结构和功能,为理解整个沉积物岩心中微生物氮的损失提供精确的信息。

图文摘要
四个采样点的底部水的地球化学和物理特征如表S2所示,采样点的水深范围为7.50至19.70 m。夏季,大亚湾底部水的盐度在28.60‰至32.50‰之间,冬季在33.10‰至33.90‰之间。夏季,底部水温范围为24.70至29.80 ,平均值为27.60,而冬季则为16.30至19.10,平均值为17.70。两个季节的pH值均在8.02至8.33之间。大亚湾夏季和冬季收集的沉积物岩心的理化参数如表1所示。夏季,NH4+含量分别为2.73±1.11至41.99 mg kg-1;冬季为2.35±0.56至10.35±1.88 mg kg-1。夏季的平均含量明显高于冬季(p <0.05)。夏季(2.73±1.41至39.63±1.78 mg kg-1)和冬季(2.59±0.12至8.32±0.77 mg kg-1)的NO3-浓度差异很大,夏季的平均值(8.45±10.01 mg kg-1)比冬季(5.15±1.62 mg kg-1)高。与NH4+和NO3-相比,两个季节的NO2-含量均较低。夏季NO2-的平均含量(0.35±0.38 mg kg-1)略高于冬季(0.23±0.17 mg kg-1)。夏季的TN含量范围为295.85±1.54至1618.46±10.54 mg kg-1,而冬季则为483.92±5.23至702.92±8.32 mg kg-1,其相对较大值出现在岩心S1(海湾内部)。夏季TN的平均含量(852.15±384.58mg kg-1)明显高于冬季(599.65±74.35 mg kg-1,p <0.05)。夏季,C含量(%)从0.61±0.01变化到1.57±0.03;冬季从0.67±0.02变化到1.53±0.04,并且未观察到C(%)的季节性变化(p> 0.05)。冬季C:N的平均值(18.47±7.02)高于夏季(13.90±3.46)。夏季,ORP值在−74.00和−324.00 mV之间;冬季在−145.00和−312.00 mV之间变化。
为了研究理化参数的垂直分布模式,用夏季每隔4 cm采集的子样本来分析理化参数沿深度的变化(图1a–g,以及表S3至S6)。NO2-的浓度(0.43±0.49 mg kg-1,n = 80)在不同岩心的不同深度处达到峰值,在S13岩心深50 cm处的最大浓度为2.41±0.02 mg kg-1(图1a)。NO3-的浓度在整个沉积物岩心中保持稳定(8.00±6.47 mg kg-1,n = 80),岩心S1除外。其中NO3-的含量随着深度的增加而逐渐增加(r = 0.808,p <0.001,n = 21),并在深度为88 cm处达到峰值(39.63±1.78 mg kg-1,图1b)。NH4+的分布在任何沉积物岩心中均无显著变化(图1c)。在岩心S5的2–6 cm深度处观察到的NH4+浓度最高(41.99±0.91至37.69±0.88 mg kg-1),在岩心S5内的NH4+浓度(14.83±9.00 mg kg-1,n = 21)高于其他岩心(5.26±1.59 mg kg-1,n = 59)。除沉积物岩心S5以外,总氮的浓度随深度逐渐降低(rS1 = − 0.677,pS1 <0.001,nS1 = 22;rS8 = − 0.872,pS8 <0.001,nS8 = 14;rS13 = − 0.889,pS13 <0.001,nS13 = 23图1d)。C(%)含量随深度的变化无明显变化(图1e)。从S1到S8站点,沉积物中TN和C(%)的含量呈递减的梯度,并且在S1岩心中检测到两者的最高浓度为1179.07±156.55 mg kg-1(n = 22)。C:N值从表层增加到40 cm深度处,然后在44 cm以下降低(图1f)。在所有样本中,沿沉积物剖面(<100.00 mV)ORP的含量降低,在岩心S1中检测到最低值(-310.21±32.18mV,n = 19;图1g)。
图1. 大亚湾沉积物中(a)NO2-,(b)NO3-,(c)NH4+,(d)TN(e)C(%),(f)C:N和(g)ORP的垂直分布。误差棒表示三份样本的标准差。据计算,夏季厌氧氨氧化的潜在发生速率范围为0.08±0.02至0.99±0.11 μmol N kg-1 h-1(图2a–d,蓝色水平柱),冬季为0.02±0.01至0.57±0.18μmol N kg-1 h-1(图2e–h)。在S1M-S(其中S表示夏季)和S1B-W(其中W表示冬季)中检测到最低的厌氧氨氧化速率,而在S5M-S和S8S-W上则观察到最高的厌氧氨氧化速率。表层沉积物中厌氧氨氧化速率(0.45±0.20 μmol N kg-1 h-1,n = 8)高于中间沉积物(0.27±0.33μmol N kg-1 h-1,n = 7)和底部沉积物(0.14±0.12 μmol N kg-1 h-1,n = 6)。表层和底部沉积物的厌氧氨氧化速率之间存在统计学上的显著差异(p = 0.021,n = 16;表S7)。总体而言,随着沉积物深度的增加,厌氧氨氧化速率有明显的下降趋势(r = − 0.463,p <0.001,n = 21)。根据潜在速率,夏季厌氧氨化菌对氮损失的贡献度为4.34%–83.53%,冬季的贡献度为10.61%–89.50%。在底部沉积物中观察到厌氧氨氧化对氮损失的最大相对贡献度(60.95±14.43%,n = 6)。厌氧氨氧化速率与NO2-的浓度呈正相关(r = 0.56,p <0.05),与深度呈负相关(r =− 0.45,p <0.05)(图5和表S10)。潜在反硝化速率高于厌氧氨氧化速率,夏季速率值的范围为0.02±0.01至6.50±1.00 μmol N kg-1h-1(图2a–d,红色水平柱)0.01±0.01至4.79±1.20 μmol N kg-1 h-1(图2e–h)。样本S1S–S和S8S–W的反硝化速率最高,而样本S1B–S和S1B–W的反硝化速率最低。与中部沉积物(0.60±0.76 μmol N kg-1 h-1,n = 7)和底部沉积物(0.11±0.14μmol N kg-1 h-1,n = 6)相比,表层沉积物的反硝化速率最高(3.31±2.04μmol N kg-1 h-1,n = 8)。表层和中部沉积物的反硝化速率(p <0.001,n = 15)以及表层和底部沉积物的反硝化速率(p <0.001,n = 14;表S7)在统计学上有显著差异。反硝化速率通常随着沉积物深度的增加而降低(r = − 0.698,p <0.001,n = 21)。在这项研究中未观察到氮损失率的季节性明显差异。根据潜在的速率,反硝化作用导致夏季氮素损失为16.47%–95.47%,冬季为10.50%–89.39%。表层沉积物中,反硝化对氮损失的相对贡献度最大(82.76±12.10%,n = 8)。反硝化速率与厌氧氧化速率呈正相关(r = 0.48,p <0.05),与深度呈负相关(r = − 0.66,p <0.01)(图5和表S10)。通过反硝化和厌氧氨氧化法估算了大亚湾沉积物中的N损失通量,如表S8所示。由于反硝化作用而损失的氮约为1.17×108t N yr-1,而由于厌氧氨氧化作用损失的氮约为3.88×107 t N yr-1,表明反硝化作用是大亚湾沉积物中微生物去除氮的主要因素。表面,中部和底部沉积物的总氮损失通量分别约为7.41×107(47.47%),6.51×107(41.67%)和1.70×107 t N yr-1(10.86%)。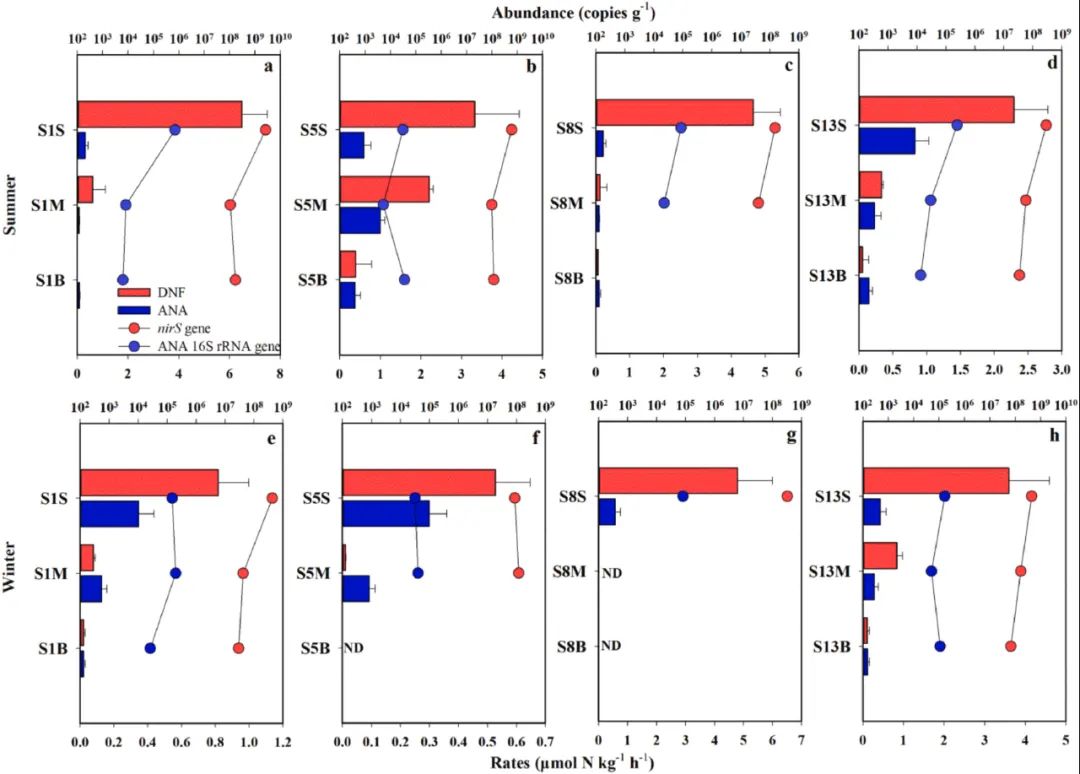
图
2. 2015年8月(a–d)和2015年12月(e–h)的大亚湾沉积物中反硝化速率及反硝化菌丰度(DNF,红色)和厌氧氨氧化速率及厌氧氨氧化菌的丰度(ANA,蓝色)。字母“S”,“M”和“B”分别代表从表面,中部和底部沉积物中收集的样本。柱状图表示沉积物中反硝化和厌氧氨氧化的速率。点代表大量的反硝化细菌和厌氧细菌。误差棒表示三份样品的标准误差。ND表示未检测到(样品不足)。通过qPCR定量的方法,分别使用nirS基因和厌氧氨氧化菌特异性16S rRNA基因估计反硝化菌和厌氧氨氧化菌的丰度。反硝化菌的丰度范围为(3.47±0.65)×107至(2.62±0.01)×109拷贝/克(夏季;图2a–d,红点)和(2.96±0.01)×107至(4.27±0.01)×108份/克(冬季;图2e–h)。样本S1S–S反硝化菌的丰度最高((2.62±0.01)×109拷贝/克),而样本S1B–S((2.96±0.01)×107拷贝/克)的丰度最低。表层沉积物中nirS基因的丰度((6.19±8.26)×108拷贝/克,n = 8)高于中间沉积物((9.04±4.65)×107拷贝/克,n = 7)和底部沉积物层((8.45±6.02)×107拷贝/克,n = 6)。随着沉积物深度的增加,nirS基因的丰度逐渐降低(r = − 0.637,p = 0.002,n = 20)。nirS基因的丰度与TN(r= 0.52,p <0.05),反硝化速率(r = 0.71,p <0.01)和厌氧氨氧化菌的丰度(r = 0.91,p <0.01)呈正相关(图5和表S10)。与nirS基因相比,厌氧氨氧化菌特异性16S rRNA基因的丰度在(5.18±0.90)×103至(7.12±0.01)×105拷贝/克之间(夏季;图2a–d,蓝点)和(2.64±0.01)×104至(1.99±0.01)×105拷贝/克(冬季;图2e–h)。表层沉积物中,厌氧氨氧化菌特异性16S rRNA基因的丰度比底部沉积物中高((1.88±2.24)×105拷贝/克,n = 8)。厌氧氨氧化菌的丰度与沉积物深度之间呈负相关(r = − 0.512,p = 0.021,n = 20)。厌氧氨氧化菌特异性16S rRNA基因的丰度与TN(r = 0.49,p <0.05)和反硝化速率(r = 0.62,p <0.01)呈正相关(图5和表S10)。在所有沉积物样本中,nirS基因的丰度比厌氧氨氧化菌特异性16S rRNA的丰度高2–3个数量级。
图5. 反硝化速率(DNF rate)、nirS丰度、厌氧氨氧化速率(ANA rate)、16S rRNA基因丰度和沉积物环境特征(包括深度、NH4+、NO3-、NO2-、TN、C(%)、和C:N)。蓝色和红色分别表示正相关和负相关。5 反硝化基因nirS和厌氧氨氧化菌特异性16SrRNA基因序列的多样性共对10万个反硝化菌和厌氧氨氧化菌的原始序列(每个样本5000个序列,n = 20)进行了去噪和修整。经过一系列质控后,筛选出3674±586个reads(n = 20)的nirS基因序列和3659±852个reads(n = 20)的厌氧氨氧化菌特异性16S rRNA序列,进行下一步分析(表S9)。基于0.03距离水平,共获得1656个nirS OTUs(每个样本407±111个OTU,n = 20)(表S9)。其中,表层沉积物中OTUs数量,Chao1和ACE指数以及Shannon指数(每个样本500±68 OTUs;620.62±58.67;718.17±79.38;4.92±0.46; n = 8)最高。nirS基因的多样性和丰富度随沉积物深度的增加而降低(OTU:r = -0.684,p <0.001;Chao1:r = -0.684,p <0.001;ACE:r = -0.720,p <0.001;Shannon:r = − 0.630,p <0.001;n = 20)。nirS基因的均匀度和覆盖度分别在0.30至0.87和0.91至0.96范围内。与nirS基因相比,厌氧氨氧化特异性16S rRNA基因共检测到340个OTUs(每个样本75±15 OTUs,n = 20)的(表S9)。中部沉积物中的OTUs,Chao1和ACE值(每个样本84±13 OTUs;100.83±17.51;138.58±24.52;n = 7)最高。Shannon指数表明,底部沉积物中厌氧氨氧化菌多样性最高的(2.56±0.56,n = 5)。厌氧氨氧化菌特异性16S rRNA基因的多样性和丰富度随沉积物深度的增加而增加(OTU:r = 0.545,p = 0.012;Shannon:r = 0.487,p = 0.029;n = 20)。此外,每个样本的均匀度和覆盖度值分别为0.27至0.73和0.98至0.99。稀释曲线分析表明,反硝化菌(图S2a)和厌氧氨氧化菌(图S2b)在当前研究中得到了很好的体现。反硝化菌(图3a)和厌氧氨氧化菌(图4a)热图分析中分别包括了50个和10个主要的OTUs。OTUs的代表性序列在系统发育上与nirS基因(图3b)和厌氧氨氧化菌特异性16S rRNA基因(图4b)进行了系统比较,这些序列主要存在于细菌分离物中(有几个来自不同采样点的未鉴定序列)。对主要的OTUs进行了系统发育分析,反硝化菌属于10个集群(图3a,b和S3)。在大亚湾的沉积物中检测到较高的反硝化细菌生物多样性。在这10个集群中,集群1-6与变形菌门的γ分支相关,而其他四个集群与变形菌门的α分支密切相关。集群6几乎在所有样品中都含有最大数量的主要反硝化OTUs,并且与Alteromonadaceae、Marinobacter和Marinobacter aquaeole密切相关。与其他样本(45±2 OTUs,n = 20)相比,S5S–S群落的主要反硝化OTUs数量最多(48个OTUs),而S1M-W和S1B–W中最少(40个OTUs)。一半的主要OTUs普遍富集在20个沉积物样本中。表层沉积物中的主要反硝化OTUs的数量较多(46 ± 1 OTUs,n = 8)。在表层沉积物下方,观察到13个相对丰度较高的OTUs(值 > 10,n = 13)。此外,在底部沉积物中,集群1-3(属于Arenimonas donghaensis)和集群8-10(未鉴定的细菌)的相对丰富的OTUs较低。PCoA分析揭示了大亚湾中反硝化菌明显的垂直分布。结果表明,表层反硝化菌群的组合与其他组合不同(34.77%的方差解释度,图3c)。冗余分析(RDA)的前两个轴(RDA1和RDA2)解释了大亚湾中反硝化细菌群落累积方差的28.44%(图S4a),且深度和亚硝酸盐是影响反硝化菌群组成的关键因素。厌氧氨氧化菌优势OTUs和GenBank中相关序列的系统发育分析表明,大亚湾沉积物中获得的所有序列属于Ca. Scalindua和四个未知集群(集群1-4,图4a和b以及S5)。共有3种优势厌氧氨氧化细菌OTUs(OTU01、OTU2和OTU08)与Ca. Scalindua有关,且普遍富集在所有样本中。其他四个集群属于来自潮间带沼泽、巴尔的摩内港、中国海岸、下北半岛和东海沉积物中的一种厌氧氨氧化菌。Ca. Scalindua在表层沉积物中占主导地位,OTU的相对丰度为2.96 ± 3.70 (n = 24)。底部沉积物中观察到未知集群(集群1-4)的OTU相对丰度较高。大亚湾沉积物中没有发现反硝化和厌氧氨氧化菌组合的时间分布。厌氧氨氧化菌群的空间变异解释了48.37%的变异。结果还表明,表层厌氧氨氧化细菌的组合与其他不同(图4c)。RDA分析(图 S4b)表明,前两个RDA轴解释了厌氧氨氧化菌与环境因素累积方差的38.66%。深度和NH4+的含量对厌氧氨氧化菌群有显著影响。
图3. 大亚湾沉积物中反硝化nirS基因的微生物特征。(a)最丰富的反硝化OTUs(前50个OTUs)的热图。(b)主要的(前50个OTUs)反硝化菌OTUs的系统发育树和来自Genbank的参考序列,Bootstrap值为1000次重复。(c)沉积物中提取的反硝化nirS基因序列的主坐标(PCoA)分析。天蓝色圆圈代表的表层反硝化菌微生物群落。红色、绿色和蓝色分别代表表层、中部和底部沉积物圆形、正方形、倒三角形和菱形分别表示S1、S5、S8和 S13的样本。夏季和冬季采集的样本分别被标记为“S”和“W”。数字后的字母“S”、“M”和“B”分别代表从表层,中部和底部沉积物中收集的样本。
图4. 大亚湾沉积物中厌氧氨氧化菌特异性16S rRNA基因的微生物特征。(a)最丰富的厌氧氨氧化菌OTUs(前10个OTUs)的热图。(b)主要的(前10个OTUs)厌氧氨氧化菌OTUs的系统发育树和来自Genbank的参考序列。Bootstrap值为1000次重复。(c)沉积物中提取的厌氧氨氧化特异性16S rRNA 基因序列的主坐标(PCoA)分析。天蓝色圆圈代表的表层厌氧氨氧化菌微生物群落。红色、绿色和蓝色分别代表表层、中部和底部沉积物圆形、正方形、倒三角形和菱形分别表示S1、S5、S8和 S13的样本。夏季和冬季采集的样本分别被标记为“S”和“W”。数字后的字母“S”、“M”和“B”分别代表从表层,中部和底部沉积物中收集的样本。沉积物是已知的氮损失的热点区,可以减少沿海水生生态系统中的人为氮输入和平衡氮收支。林等人发现东海表层沉积物是去除人为溶解性无机氮输入重要的汇,反硝化去除氮约为2.20 × 106 t N yr−1,厌氧氨氧化去除氮约为4.60 × 105 t N yr−1。然而,沿沉积物深度梯度从亚氧界面到缺氧底部沉积物对氮损失的贡献是有限的。根据反硝化和厌氧氨氧化的平均潜在速率,地表(S)、中部(M)和底部(B)沉积物中的年氮损失率根据等式4(表S6)进行估算。结果表明,表层、中部和底部沉积物每年分别去除7.41×107 t(47.47%)、6.51×107t(41.67%)和1.70×107 t(10.86%)的氮。表层沉积物中反硝化年通量(6.53 × 107 t N yr−1)高于厌氧氨氧化(8.87 × 106 t N yr−1),而底部沉积物中则相反,反硝化和厌氧氨氧化年通量分别为7.49 × 106 t N yr−1和9.47 × 106 t N yr−1。因此,表下沉积物的年氮损失率与表层沉积物相等。沿大亚湾的沉积物岩心深度(0-90厘米深)发现反硝化细菌和厌氧氨氧化细菌的潜在活动。表层沉积物中,反硝化作用是氮损失的主要途径(82.76 ±12.10%,3.31 ± 2.04μmol N kg−1 h−1, n = 8,图6a和b),低于珠江入海口、长江入海口、胶州湾和中国沿海湿地的表层沉积物。大亚湾表层沉积物的厌氧氨氧化速率(0.45±0.20μmol N kg−1 h−1,n = 8,图6a和b)与新河入海口、九龙湖水库和中国河口沉积物报道的值一致。随沉积物深度观察到不同的反硝化和厌氧氨氧化速率的垂直分布模式。在表层以下,反硝化和厌氧氨氧化速率逐渐降低(p < 0.001,n = 16),正如我们在之前的研究中所预期的那样。Seitzinger研究表明,沉积物孔隙水中硝酸盐的有效性通常是控制反硝化速率的关键因素。本研究中TN浓度随沉积物深度而降低(图1e),表下沉积物中无机氮(如亚硝酸盐和硝酸盐)含量低于表层沉积物。这些结果表明反硝化作用受无机氮有效性的调节。与沉积物岩心中的亚硝酸盐浓度相比,本研究中铵含量不受厌氧氨氧化活性的限制。厌氧氨氧化速率与NO2-呈正相关(r = 0.56,p <0.05),与深度呈负相关(r = - 0.45,p <0.05)。亚硝酸盐含量可能是厌氧氨氧化活性的关键控制因素。综上所述,电子受体中亚硝酸盐和或硝酸盐的有效性可能是影响氮损失过程的重要因素。此外,以往的研究还发现,氧通常被认为是影响沉积物中氮损失过程的重要因素。但由于氧气渗透有限,氧气水平只会影响最上层的表层沉积物(<1 cm)。ORP值在表层沉积物以下变化不大,且值较低(<100.00 mV)。因此,反硝化和厌氧氨氧化似乎受沉积物岩心氧含量的影响较小。更重要的是,厌氧氨氧化的相对贡献度随着沉积物深度的增加而增加,并且是导致表层沉积物以下氮损失的主要过程(底部沉积物(90 cm深度)中高达60.95 ± 14.43%,n = 6;图6a和b)。该值明显高于沿海沉积物的文献值和全球生态系统的平均水平(23%)。厌氧氨氧化对氮损失的相对贡献度随沉积物深度增加的趋势表明,反硝化速率比厌氧氨氧化速率下降的更为显著。在深层旱地土壤中也有类似的结果,氮的损失主要由低于20厘米的厌氧氨氧化过程引起(夏季65.40±12.50%;冬季79.40±14.30%)。反硝化是一个依赖于有机物消耗的异养过程,而厌氧氨氧化是一个自养过程。研究发现表层沉积物中含有大量不稳定有机质,而顽固性腐殖质是沉积物中的主要成分。因此,缺乏不稳定有机物会削弱反硝化作用并加强厌氧氨氧化。在沉积物岩心中未观察到反硝化作用和厌氧氨氧化速率的季节性变化,表明季节变化引起的环境参数,包括温度、盐度、原位水文条件,不能显著影响大亚湾沉积物岩心的氮损失。与海水相比,由于沉积物的固体基质,深层沉积物不受环境特征季节性变化的影响。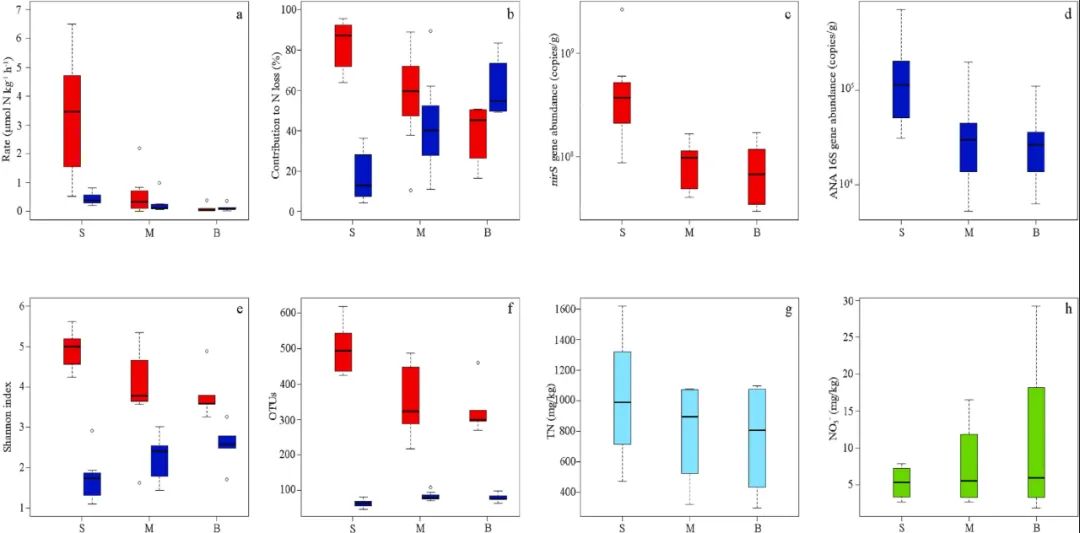
图6. 沉积物岩心反硝化及厌氧氨氧化特性及关键理化参数的深度分布(a)反硝化和厌氧氨氧化速率的垂直分布;(b)对氮损失的贡献度;(c)nirS型反硝化菌的丰度;(d)厌氧氨氧化菌的丰度;(e)基于OTUs的反硝化和厌氧氨氧化菌Shannon指数;(f)反硝化菌和厌氧氨氧化菌的OTUs;(g)TN浓度的深度分布;(h)硝酸盐含量的深度分布。在箱线图中,中间线代表中值,垂直线代表最大值和最小值。圆点表示异常值,红色和蓝色框表示反硝化和厌氧氨氧化过程。大写的S、M、B分别代表沉积物核心的表层(S)、中层(M)和底层(B)。大亚湾表层沉积物中nirS基因的平均丰度为6.19×108拷贝/克(n = 8,图6c),相对高于切萨皮克湾、旧金山湾、莱州湾、长江入海口、南海、黄河入海口沉积物和稻田土中观察到的丰度。在所有样本中,厌氧氨氧化菌特异性16S rRNA 基因的丰度比nirS基因低2-3个数量级(图6d),也低于开普菲尔河入海口、长江入海口、珠江入海口沉积物、稻田土壤以及中国的沿海湿地。此外,反硝化菌和厌氧氨氧化菌的丰度垂直分布不同。底部沉积物中,nirS基因的基因拷贝数下降至(8.45 ± 6.02) × 107拷贝/克(n = 5,图6c),而厌氧氨氧化酶丰度下降至(3.85 ± 4.17) × 104拷贝/克(n = 5,图6d)。在田间土壤和城市湿地中发现随着沉积物深度的增加,反硝化菌和厌氧氨氧化菌的丰度也出现类似的下降。沈等人发现浅层沉积物是厌氧氨氧化菌生长的首选栖息地。沿海沉积物岩心的地球化学特征可能是降低反硝化菌和厌氧氨氧化菌丰度的关键因素。在本研究中,nirS基因(r = 0.52,p < 0.05)和厌氧氨氧化菌特异性16S rRNA基因(r = 0.91,p < 0.01)的丰度与TN含量呈正相关。除DO和有机碳外,TN含量的有效性可能是控制表下沉积物中反硝化和厌氧氨氧化细菌丰度的关键因素与珠江入海口沉积物岩心的研究结果相似,大亚湾沉积物岩心中未观察到nirS型反硝化菌和厌氧氨氧化菌的显著季节性差异。与克隆文库的数据相比,对nirS基因和厌氧氨氧化菌的16s rRNA基因的 MiSeq高通量测序可以提供足够的信息来揭示群落变异,甚至可以检测到相对丰度较低的稀有微生物。基于OTUs和Shannon指数,表层沉积物中的nirS基因说明了高度多样化的反硝化菌群落(图6e和f)。反硝化菌的Shannon指数和OTUs随沉积物深度的增加而降低,底部沉积物中的值最低(Shannon指数和OTUs分别为 3.82±0.63和330.00±75.00)。陈等人还发现反硝化菌的多样性随土壤深度的增加而下降。在珠江入海口也发现了类似的反硝化菌下降的趋势。较高的不稳定有机物和氮含量可能有利于各种反硝化菌的生长。相比之下,厌氧氨氧化菌的丰富度和多样性随着沉积物深度的增加而增加(图6e和f),表明底部沉积物具有稳定的缺氧和低竞争环境,是厌氧氨氧化菌的合适栖息地。在70-80厘米深度的菜地中也发现了较高的厌氧氨氧化菌多样性(Shannon指数为2.585)。反硝化菌的主要OTUs属于变形菌门的α和γ分支,且珠江入海口的沉积物岩心处也具有10个集群的高度多样性。本研究中,集群1-6与Alteromonadaceae、Marinobacter和Marinobacter aquaeoolei密切相关(图3)。谢等人发现反硝化菌的主要OTUs属于变形菌门的α、β和γ分支。一些研究表明,系统发育多样化的微生物能够执行反硝化作用,并且大多数先前描述的反硝化菌属于变形菌门的亚门。此外,大亚湾表层沉积物中也发现了δ-变形菌纲和放线菌纲。未观察到沉积物岩心中反硝化菌群组成的季节性变化。高等人指出,反硝化菌的分布反映了对场地特定特征的适应。但是,PCoA分析表明,表层反硝化菌的组合与其他组合不同(图 3c)。在表层沉积物之下,反硝化菌的群落组成总体上随着沉积物深度的增加而变化较小,除非出现了几个相对丰度较高的OTUs(值 > 10,n = 13)(图4)。优势OTUs的冗余分析(RDA)表明深度和亚硝酸盐是影响反硝化菌群落组成的关键因素。类似于海洋环境中Ca. Scalindua,在大亚湾中总共发现了三种与Ca. Scalindua密切相关的厌氧氨氧化细菌OTUs(OTU01、OTU02和OTU08),且在表层沉积物中占主导地位,相对丰度为2.96 ± 3.70(n = 24)。Ca. Scalindua rubra的草图基因组分析表明其能够适应高盐的环境。最近,吴等人通过高通量测序分析发现,Ca. Scalindua仍然是海洋沉积物中的优势属,但不是唯一的属。这表明引物和测序深度将直接影响厌氧氨氧化菌的群落组成分析。本研究采用引物组 Amx368F和Amx820R分析河口和海洋生态系统中厌氧氨氧化菌的分布和多样性,具有良好的覆盖度。PCoA分析结果表明表明表层厌氧氨氧化菌的组合与其他组合相分离(图4c)。表层沉积物以下的深处观察到未知集群(集群1-4)的相对丰度较高,这表明新集群可能是由PCR扩增偏差或由未鉴定的厌氧氨氧化菌的真实存在引起的。根据RDA分析(图S4b),深度和NH4+对厌氧氨氧化菌群有显著影响。NH4+浓度一直被看作是塑造沉积物中厌氧氨氧化菌群结构的关键因素。表层沉积物是微生物氮转化最活跃的栖息地,具有较高的潜在氮损失速率,尤其是反硝化速率。表层沉积物中,来自水体的下沉有机物(电子供体)通过氧化提供能量,直接或间接驱动氮损失过程。将NO3-还原为N2的反硝化菌依赖于消耗有机物,而自养厌氧氨氧化菌将NH4+和NO2-结合产生N2。因此,表层沉积物中的反硝化速率受到刺激,在氮损失过程中占主导地位(80%)(图6a-b)。异养反硝化细菌的多样化群落是由于水中更富含能量的底物的更高可用性而得以维持。相比之下,厌氧氨氧化菌无法在富含有机物的环境中与反硝化菌竞争亚硝酸盐,因此厌氧氨氧化速率和丰度低于表层沉积物中的反硝化菌(图6a)。此前的研究表明,表层沉积物在富氧水中不断处于悬浮状态,使有机物迅速降解。表层沉积物中含有更高浓度的NH4+(图S6b),表明大多数异养细菌进行的氨化作用在含氧表层沉积物中更强。同时,表层沉积物的亚硝酸盐和硝酸盐浓度较低(图 6h和S6c),这意味着亚硝酸盐或硝酸盐通过耦合硝化反硝化或厌氧氨氧化转化为N2。先前的研究表明海洋沉积物中稳定的好氧/缺氧界面为硝化和反硝化之间的耦合提供了完美的条件。硝化菌和厌氧氨氧化菌位于相同的栖息地这一事实近一步证实了硝化和厌氧氨氧化耦合的假设。因此,耦合硝化反硝化或厌氧氨氧化在大亚湾表层沉积物中氮的去除起重要作用。然而,随着沉积物深度的增加,沉积物中几乎所有的溶解氧都被消耗掉,表层沉积物中溶解性无机氮和有机物质的扩散和平流输送受到限制(图1、6g和 6h)。难降解有机物占海洋沉积物中TOC的72-97%。源自海洋浮游植物的有机质沉积在沉积物表面,转变为腐殖质或难降解的有机物,然后沉入沉积物底部。难降解有机物含量随沉积深度增加而增加。大部分不稳定有机物主要被表层沉积物中的好氧和厌氧微生物过程消耗。表层沉积物中剩余的大部分难降解有机碳和少量未消耗的不稳定有机碳将继续埋藏在深层沉积物中。因此,可用于反硝化的电子供体在底部沉积物中受到限制,也限制了深层沉积物中的反硝化活性。然而,深层沉积物中有机物的矿化和表层沉积物中未消耗的不稳定有机碳的下沉可以为反硝化提供有限的电子。依靠这种不稳定的有机碳,在深层沉积物中仍然可以起到一定的反硝化作用。如图2所示,在所有底部样品中都检测到反硝化率,表明表下沉积物在氮去除中的作用不应被忽视。本研究中,表下沉积物中较低的反硝化速率、多样性和OTUs(图6a、e和 6f),可能是由于富含能量的底物的有效性较低所致。厌氧氨氧化菌的多样性(Shannon指数和OTUs)高于表下沉积物中的反硝化菌。厌氧氨氧化速率与NO2-呈正相关(r = 0.563,p < 0.05),而铵是影响厌氧氨氧化菌组成的关键因素。NO2-浓度在20-80 cm深度较高,这为表下沉积物中的厌氧氨氧化菌提供了NO2-。由于底部沉积物中较高的潜在NO2-源较高且有机物含量较低,与表下沉积物中的反硝化作用相比,厌氧氨氧化主导了氮损失过程。表下沉积物中NH4+和NO2-的浓度较低(图S6b和S6c),表明氨化和硝化速率较低。尽管如此,NO3-浓度随着沉积物深度而增加(图6h),表明NH4+氧化为NO2-/NO3-可能是在缺氧条件下发生的。有证据表明,缺氧硝化作用,包括NH4+与铁氧化物、锰氧化物或硫酸盐的反应,可能发生在更深的缺氧沉积物中,以提供可用的溶解性无机氮。厌氧硝化可以提供地下沉积物中反硝化和厌氧氨氧化氮损失所需的NO2-或NO3-。因此,耦合的缺氧硝化和厌氧氨氧化或反硝化在大亚湾中的表下沉积物中占主导地位。本研究强调了沿海海湾沉积物岩心中氮损失的贡献者。根据对每年氮损失的估计,表下沉积物的作用与大亚湾中的表层沉积物几乎相等。反硝化作用是表层沉积物中主要的氮损失过程(>80%),而厌氧氨氧化作用强于底部沉积物的反硝化作用(>60%)。反硝化和厌氧氨氧化的潜在速率和基因丰度都随着沉积物深度的增加而降低,但厌氧氨氧化菌的多样性和丰富度随沉积物深度的增加而增加。硝化和反硝化或厌氧氨氧化的耦合对去除表层沉积物中不能完全去除的埋藏或运输氮具有很高的潜在贡献度,从而保持底部沉积物中较低的溶解性无机氮水平。








































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










