编译:微科盟傻狍子,编辑:微科盟木木夕、江舜尧
微科盟原创微文,欢迎转发转载,转载须注明来源《微生态》公众号。
导读
了解陆地生态系统对于应对全球气候变化具有重要意义。但是,大多数实验方法都集中在单一因素上。在自然系统中,湿度和温度经常同时变化,它们可以相互作用并影响微生物的响应。尽管土壤湿度和温度是控制微生物活动的非常重要的因素,但是当两个变量同时变化时,微生物生长速率对温度和湿度的依赖性及其敏感性仍然存在差异。本实验创建了一个湿度梯度,并确定了温度对细菌和真菌生长率以及呼吸率的依赖性。结果表明,微生物速率(microbial rates)随着湿度的降低而降低,而随着温度的升高而升高,直至达到最大值;并且还发现微生物生长和呼吸的温度敏感性(Q10)不受湿度变化的影响。这些结果为验证生态系统和全球循环模型(GCMs)中使用的温度和湿度变化假设提供了一个实验框架。
原名:Can moisture affect temperature dependences of microbial growth and respiration?
译名:湿度会影响微生物生长和呼吸对温度的依赖性吗?
期刊:Soil Biology and Biochemistry
IF:7.609
发表时间:2021.03
通讯作者:C. Cruz-Paredes
通讯作者单位:隆德大学生物系
土壤样本取自瑞典北部阿比斯科研究站附近的亚北极白桦林,土壤为有机土。样本取自4个距离不小于10 m的随机分配样地,以获取研究系统内的空间变异性。土壤有机质(SOM)采用着火损失(LOI)法测量。土壤全碳(C)和全氮(N)采用Dumas干燃烧法,使用C/N 144元素分析仪进行分析。采用均质/离心技术,将3H-亮氨酸掺入细菌中,在0.5 g土壤中测量细菌生长。提取细菌中的亮氨酸含量作为细菌生长的代理指标。在0.5 g土壤中使用14C-醋酸酯转化为真菌特异性脂质麦角甾醇测定真菌生长。提取麦角甾醇中乙酸酯的加入量作为真菌生长的代理指标。为进行呼吸测量,将0.5 g的土壤称重到20毫升的玻璃小瓶中,用压缩空气净化并密封。通过配备甲烷化器和火焰电离检测器的气相色谱仪来测量二氧化碳。根据不同温度调整了培养时间,以确保C的使用水平相似。微生物CUE由微生物生长(细菌生长和真菌生长之和)除以微生物C碳使用量(呼吸和微生物生长之和)估算。微生物生长和呼吸对水分的依赖性采用下式模拟:

其中,y为过程速率,x为土壤湿度,a为所测过程受50% (IC50)抑制时的土壤湿度,c为WHC为50%时的最优过程速率,b为抑制率参数。为了发现温度对水分依赖是否有任何影响,估算了每种温度下细菌生长、真菌生长和呼吸的IC50值,并根据温度进行回归。根据公式(2),使用Ratkowsky模型模拟微生物生长的温度依赖性:

在低于最佳温度时,呼吸速率可以使用简化的方程表示:
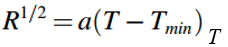
T为筛选温度(℃),Tmin为最低温度。温度系数(Q10)是温度系数(Q10)随温度每10℃的变化而变化的速率。根据下式估算出Q10作为温度敏感性指标:

采用单因素方差分析,以10、20、30和50% WHC 4个水平的土壤湿度为因子,检验不同速率下Tmin、Topt、Tmax和Q10的差异。此外,当发现任何微生物速率存在显著差异时,进行线性回归,以检验湿度水平造成的系统差异。
土壤有机质含量变化范围为51~70%(表1)。土壤pH值为酸性,介于4.4~4.9之间。电导率在51.5-145.5之间变化。C/N比值在34.1~39.1之间变化(表1)。
在所有温度下,土壤湿度越低,细菌的生长速度越慢(图1A)。同样,在所有温度下,土壤湿度越低,真菌生长速度越慢,但其坡度比细菌生长的坡度要小(图1A)。此外,在实验中包含的最低水分条件下,细菌的生长速度约为50% WHC时的1/10,而真菌的生长速度仅为50%。除45℃外,在所有温度下,呼吸速率与微生物生长速率呈相同的趋势,并随土壤湿度的降低而降低(图1B)。将微生物生长速率和呼吸速率转化为C单位,以估算CUE(表2)和真菌生长速度。真菌生长的速度也表明,真菌对低水分的耐受性更高(图1)。与微生物的生长速率和呼吸速率相比,CUE的相对速率不受水分变化的影响(图1 B),并且在低于最佳生长温度的所有水分含量下,其CUE值均相似。微生物生长和呼吸速率的水分依赖性可以用logistic模型描述(R2值在0.85到0.99之间)。细菌和真菌生长速率的IC50分别为WHC的17-24%和7-10%。此外,微生物生长速率的IC50值不随温度变化。另一方面,呼吸作用的IC50值在8~16% WHC之间。呼吸速率的IC50值在0 - 25℃之间没有变化,但是超过30℃时,IC50值会下降到45℃,而无法估计IC50值。
图1 20℃时不同湿度水平下的微生物生长速率。A)标准化的细菌(●)和真菌(◆)生长速率和真菌占微生物生长的比例(
▴
)。B)标准化的呼吸速率(◣)、微生物生长速率(◆)、碳利用效率(CUE)值(■)和真菌占微生物生长的比例(
▴
)。点表示平均值,误差条表示标准误差。
表2 在不同的湿度下,细菌和真菌生长和呼吸速率、真菌与细菌(F/B)比率和CUE值。
微生物的生长速率随着温度的升高而增加,直至达到最佳温度(Topt),细菌的最佳温度为30℃,真菌的最佳温度为32℃(表3),然后细菌的生长速率下降直至达到最高温度(Tmax)(表3)。在0–20℃的区间内,微生物加工速率与温度呈线性关系。细菌和真菌生长对温度的依赖性相似,并且两者均可由Ratkowsky模型描述(图2A和B),所有样品的平均R2值为0.96。呼吸的平方根呈线性关系,平均R2值为0.98。,我们发现CUE常量值在0到20℃之间约为0.05,之后CUE值下降到约0.001(图2D)。在所有水分水平都是如此(图2)。但是,随着土壤湿度的降低,细菌在最佳温度下的生长和呼吸速率的幅度降低了。如上所述,细菌生长速率对水分减少的敏感性高于真菌生长速率。
图2 平方根转化率的温度依赖性:将A)细菌生长,B)真菌生长,C)呼吸速率归一化到生长的最适温度(Topt)。由于归一化,微生物的生长和呼吸速率是无单位的。拟合曲线基于Ratkowsky模型。在最佳温度以下,平方根转化率显示出对筛选温度的线性响应。D)碳使用效率(CUE)的温度依赖性。值是四次重复的平均值,误差线代表标准误差。不同的土壤水分用不同的颜色表示。

在建立了每个湿度处理的温度依赖关系后,我们估算了所有微生物过程的Tmin(图3A)和Q10(5-15℃)(图3B)值以及微生物生长的Topt和Tmax(表3)值。土壤水分水平对细菌和真菌生长速率的Tmin值无显著差异(F3,12=0.39,p=0.76;F3,12=1.95, p=0.18),但由于10和30% WHC值之间的差异,呼吸速率存在显著差异(F3,12=4.35, p=0.03)(图3A)。然而,当使用线性回归分析是否存在系统效应时,结果表明湿度变化与Tmin值之间没有显著的关系(R2=0.0003,p=0.95;R2=0.2,p=0.06;R2=0.2,p=0.06)。对于Topt和Tmax,土壤水分水平对细菌和真菌生长速率的影响均无显著差异(F3,12=2.02,p=0.17;F3,12=2.52,p=0.11;F3,12=1.5,p=0.27;F3,12=0.26,p=0.85)。实验中没有发现土壤水分水平对细菌或真菌生长速率的Q10值有任何显著影响,但对呼吸速率有显著影响,推测是10和30%WHC值之间的差异(图3B)。然而,通过线性回归分析系统效应时,湿度变化与Q10值之间没有显著的关系(R2= 0.001, p=0.89;R2=0.04, p=0.47;R2=0.18, p=0.06)。
图3 A)在不同湿度下细菌(●),真菌(◆)和呼吸(◣)速率的Tmin值和B)Q10(5-15°C)值。值代表四次重复的平均值,误差线代表标准误差。使用单因素方差分析,以土壤湿度作为四个水平的因子来检验处理差异,产生p值和R2值。
为了能够确定湿度是否改变了微生物对温度的依赖性,本实验创建了一个土壤湿度实验范围(从10%WHC到50%WHC),在此范围内,确定了细菌和真菌生长速率以及呼吸速率的温度依赖性。将不同湿度的土壤进行十种不同温度下的培养,从而确定内在的温度依赖性。大多数生态系统模型需要固有的温度依赖性来确定温度和湿度的速率调节因子,因此,在本研究中区分了直接和间接的温度依赖关系。微生物的生长速率和呼吸速率随湿度的降低而降低。这些结果与之前的研究相一致,即土壤干燥时微生物活性降低。此外,我们发现真菌对低湿度条件更有抵抗力,这也符合温暖和干燥生态系统的环境影响。研究表明,CUE值没有系统地随着湿度范围变化,因为微生物生长和呼吸速率都受到湿度的相似影响。表明在稳定的湿度条件下,不同的湿度不会影响CUE。然而,关于CUE如何响应水分变化的研究之间存在分歧。在将土壤干燥至不同水分含量的实验室实验中,发现低水分水平下CUE更高。在田间处理中,降雨间隔时间延长的土壤中CUE值更高,在旱季表现出更高的值。然而,其他研究表明,在干燥土壤中CUE下降,这可能与获取基质增加以抵御渗透冲击有关。其他研究发现,细菌的IC50值在WHC的15%到20%之间。对于呼吸速率,本研究表明当温度超过25°C时,干旱敏感性下降。由于所获得的结果与之前的发现相符,并且所选水平涵盖了微生物速率快速变化的水分含量范围,因此本实验可以验证是否包括了相关且全面的水分含量,从而证实实验设计的准确性。因此,本实验可以测试温度依赖性是否受到不同的湿度水平的影响。本实验确定了在受控土壤水分的实验范围内细菌生长,真菌生长和呼吸速率的温度依赖性。实验中对温度的依赖遵循了在以前的研究中看到的预期模式,并且由Ratkowsky模型描述。在最适温度之前(Topt),微生物的生长速率随着温度的升高而增加,在此温度之后,微生物的生长速率在Tmax时下降至零。宏观分子速率理论中提供了对温度超过最佳值时观测到的下降速率的机械解释,这是经典Arrhenius方程的理论参数化。在这一解释中,微生物活性超过最佳的降低主要归因于酶在高温下的分子完整性和酶活性的逐渐丧失,从而逐渐丧失了酶的活性。Ratkowsky模型可以很好地捕获与MMRT相同的微生物温度关系。如实验观察到并详细讨论的那样,呼吸速率在测定所包括的整个温度范围内都增加了。利用这些模型的温度依赖性曲线,我们估算了生长的最低温度(Tmin),生长的最佳温度(Topt)和温度敏感性(Q10(5-15℃))。这些温度关系的基点可以用作整个温度关系的有效指标。由于可以以最高的精度确定Tmin参数,因此它对筛选温度关系的变化是最关键的。在细菌生长中,Tmin为-9.6℃,Topt为30℃,Q10(5-15℃)为2.7。对于真菌生长中,Tmin为-8.1℃,Topt为32.8℃,Q10(5-15℃)为3.0。呼吸Tmin为-7.8°C,Q10为3.2。这些值与先前研究中有关这些指数如何根据平均年温度变化的预测相一致。在南极梯度中,细菌的Tmin为-15至-9℃,低于本研究中发现的Tmin,因为南极温度在-4至-1.5℃之间。同样,在温带森林地区,在6.5至8℃(均高于我们的研究温度)的热带地区,细菌的Tmin较高。此外,南极土壤的Q10(0-10°C)值较低,而细菌的Tmin值较低,而温带草原和温带森林地带的Q10(0-10°C)值较低。只有少数研究评估了真菌对温度的依赖性。最近在瑞典南部草原上的一项研究中,真菌的Tmin为-10℃,而在高海拔热带地区则为-6℃。这两项研究都不符合高海拔热带地区的冷适应温度关系。在本实验的研究地点中,真菌的Tmin- 8℃,表明它比在高海拔热带地点发现的真菌更适应寒冷的温度关系,但比在瑞典南部草原发现的真菌更适应温暖的温度关系。最后,在瑞典南部的两个不同研究中,呼吸Tmin的值分别为-6ºC和-5ºC,
Q10
(5-15ºC)为4,而在本研究中,Tmin为-7.8和Q10(5-15ºC)为3.2。这与预期一致,即在较冷的环境中,真菌的呼吸温度关系更适应冷(即Tmin低),这也意味着它们的温度敏感性较低。总的来说,本研究估算的微生物温度关系与先前研究的预期相符。本实验估算了C的使用效率(CUE)及其温度依赖性。CUE随温度升高而保持恒定,直到约25°C为止,此后CUE下降直至在45°C时几乎为零。这些结果与微生物生理学文献的预期相符,即当生物体暴露于接近或超过其生理极限的温度时,微生物CUE会降低。该结果也与其他的报告一致,即CUE对5℃至20℃的温度变化无响应。另外,研究表明,随着温度的升高CUE的变化很小。但是,一些研究发现土壤变暖会降低CUE,而另一些研究则发现它会增加CUE。这些分歧可能是由于采用了不同的方法来测量CUE。例如,13C方法可追踪13C标记底物的摄取,可推断出生长速率,并估算出添加底物的CUE。这种方法的结果可能会高估CUE。18O示踪法是将18O标记的水掺入DNA中以测量总生长速率,导致CUE的估计值较低。本实验采用的方法基于相似的原理,通过提供总微生物生长的估算值来追踪18O进入DNA的过程,然后通过其与微生物生长和呼吸的总和相关以估算CUE。但是,尽管CUE值通常较低,但我们可以得出结论,CUE估计值不会因湿度差异或低于Topt的温度差异而发生变化。本实验在土壤水分的实验范围内,确定了细菌生长,真菌生长和呼吸速率的温度依赖性以及温度敏感性指数。在这项研究中,发现土壤水分不会改变微生物生长或呼吸速率的内在温度关系。通过在所计算的预测变量:Tmin,Topt,Tmax或Q10(5-15ºC)在整个土壤湿度实验范围内缺乏系统的变化,可以明显地看出微生物与温度之间的关系。同样,实验中的其他研究也没有发现土壤呼吸对温度的敏感性因土壤水分的差异而发生重要变化。此外,我们还发现温度不会影响微生物对水分的依赖性或对干旱的敏感性(IC50)。这些结果表明,土壤湿度和温度独立地改变了微生物的生长和呼吸,这证实了在土壤C模型中实施的广泛使用但未经检验的假设。需要强调的是,在这项研究中,土壤的孵育时间很短,从而将混杂因素(例如底物限制)对温度依赖性的影响降到最低。与本实验的发现相反,先前的研究已经注意到湿度和温度之间的相互作用。但是,这些研究只估计了对总速率的影响,而总速率也受表观温度依赖性的影响。之前的研究表明,由于水分的影响,气候变暖对土壤呼吸的表观温度敏感性的影响随季节变化,因为底物的利用率下降并成为限制因素,导致表观Q10降低。此外,其他的研究表明,在同一草原上的两种土壤中,水分对温度敏感性的影响存在差异,其中一种土壤在中间土壤湿度下具有最高的温度敏感性,而另一种土壤的温度敏感性随着土壤湿度的增加而增加。在本研究中,实验条件涵盖了从最佳水分含量(50%WHC)到低水分含量的一系列水分,证明了选定的水分含量范围会迅速改变微生物速率。这表明本实验包括了相关且全面的水分含量范围。然而,我们没有研究高湿度条件下的厌氧状态。众所周知,在高水分含量下,微生物活动可能会出现其他限制,如氧气扩散受限。其他的研究结果表明,只有在氧气不是限制因素的情况下,温度和水分才可以在较高水平下增加分解。此外,土壤湿度只会在极端水平上影响土壤呼吸温度敏感性,而在测试的其他湿度条件下,温度敏感性不受湿度影响。因此,对于饱和土壤氧气限制普遍存在的生态系统,研究较高的湿度对微生物生长的影响可能是有意义的。作者在本文中通过改变湿度梯度,对样品中微生物生长速率进行研究。通过构建Ratkowsky模型,来比较不同湿度对微生物生长和呼吸的温度依赖性产生的影响。之前对于微生物生长的研究大多仅集中在单一因素上,而本实验同时对温度和湿度两个因素与微生物生长的关系进行研究。结果表明,在最适温度之前(Topt),微生物的生长速率随着温度的升高而增加,在此温度之后,微生物的生长速率在Tmax时下降至零,而土壤湿度对微生物速率的温度依赖性没有影响。为进一步研究温度和湿度对微生物生长和呼吸的影响提供了数据支撑。








































 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










