


















重磅综述 | Science:微生物组与人类癌症




几个世纪以来,微生物在癌症发生、诊断、预后和治疗中的作用一直存在...
导读
几个世纪以来,微生物在癌症发生、诊断、预后和治疗中的作用一直存在争议。最近的研究表明,细菌、病毒和/或真菌在癌症中普遍存在,是癌症免疫治疗的关键因素,并可用于治疗癌症转移。尽管有这些发现,已知的直接导致癌变的微生物数量仍然很少。从现代癌症生物学的角度对这些证据进行批判性的评估和建立“微生物意识”框架是一项重要任务。在这篇综述中,作者描述了微生物与癌症的因果关系和协同作用,并追踪了它们与宿主免疫系统的互作关系,此文称为免疫肿瘤-微生物轴(immuno-oncology-microbiome,IOM)。最后,作者还进一步回顾了肿瘤内微生物相关研究及操纵宿主肠道或肿瘤微生物组的方法,同时预测了下一阶段可能的发现。
论文ID
原名:The microbiome and human cancer
译名:微生物组与人类癌症
期刊:Science
IF:41.845
发表时间:2021.3
通讯作者:Rob Knight
通讯作者单位:美国加州大学圣地亚哥分校生物工程系
综述框架

主要内容


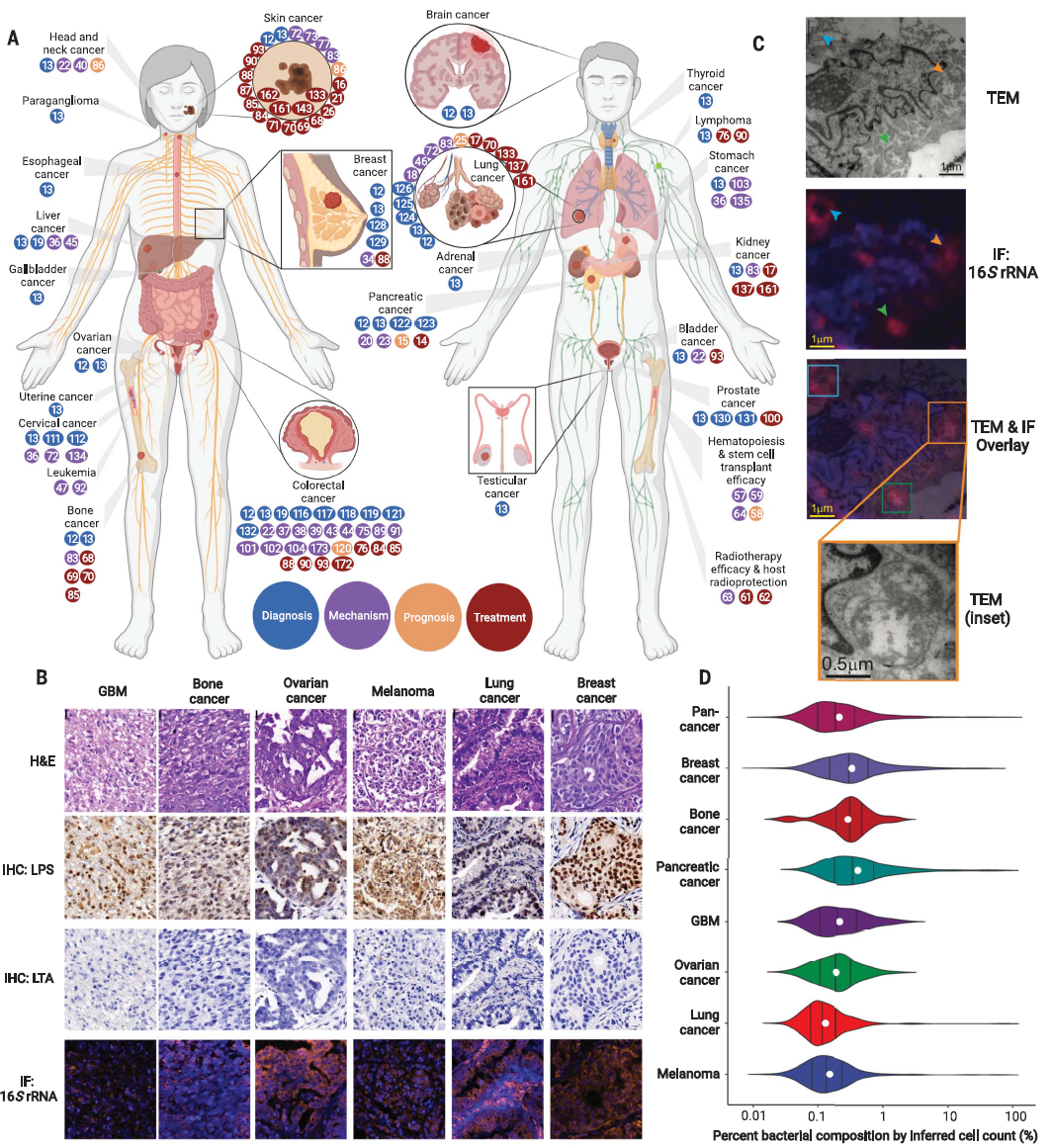


展望与讨论

评论

不感兴趣
看过了
取消
本文由“健康号”用户上传、授权发布,以上内容(含文字、图片、视频)不代表健康界立场。“健康号”系信息发布平台,仅提供信息存储服务,如有转载、侵权等任何问题,请联系健康界(jkh@hmkx.cn)处理。
关键词:
微生物,抗生素,癌症

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










