


















代谢竞争:昼夜节律稳态和肿瘤发生




我们假想分子时钟掌控肿瘤细胞的分化和其干细胞特性(即高增殖性)的转化。

哺乳动物的生物钟
环境适应能力(例如昼夜节律)对于哺乳动物内环境稳定至关重要。哺乳动物拥有不同等级的生物钟,包括下丘脑视上核(SCN)的中枢生物钟和外周生物钟。中枢生物钟感知光亮,并使组织在黑暗中保持同步的自主节律,确保生物能够适应环境和太阳周期的变化。外周生物钟对营养的摄取十分敏感,因此对细胞代谢影响巨大,摄入食物成分的变化可以导致外周生物钟的组织特异性基因表达发生改变。SCN通过控制自主神经系统和内分泌因子(如糖皮质激素和胰岛素)的分泌而与外周生物钟相互作用,从而在时间上彼此同步,最终协调新陈代谢的昼夜节律、行为和生理学。总之,复杂、时空相关、敏感通路和反映细胞能量状态的通路在染色体水平上整合到生物钟系统中(图1)。
从分子角度而言,生物钟基于转录-翻译反馈环。多个转录因子(TFs)、核受体和共调节因子通过结合特异的DNA识别序列,如E-盒、视黄酸受体相关的孤儿受体(ROR)反应元件(RRE的)和D-盒,参与驱动生物的周期性。具体而言,昼夜节律条件包括由光或胰高血糖素诱导的cAMP响应元件结合蛋白(CREB)介导的Per1产生、由糖皮质激素诱导的糖皮质激素受体(GR)介导的Per1上调、由胰岛素诱导的mTOR复合物1(mTORC1)介导的PER2产生。转录激活子昼夜运动输出周期蛋白(circadian locomotor output cycles kaput CLOCK)和脑与肌肉中的ARNT-like1蛋白(BMAL1)形成异源二聚体,并绑定到启动子E盒序列上的靶基因,驱动时钟控制基因(CCG)转录。亦由CCG 编码的昼夜节律调节器(PER)和和隐色昼夜节律调节器(CRY)蛋白在夜间抑制CLOCK–BMAL1介导的转录。Per1,Per2,Per3和Cry2的表达主要受CLOCK–BMAL1分支调控。维甲酸受体相关的孤儿受体(ROR)和REV-ERB家族成员也由CCG编码并激活或抑制Bmal1转录(激活子RORs在白天达到峰值,抑制子REV-ERBS在夜间达到峰值)。D-box结合蛋白(DBP),肝细胞白血病因子(HLF),甲状腺营养因子(TEF)和核因子,白介素3调节(NFIL3)由CCG编码并激活或抑制Rorc转录。除了这些环境信号外,核心时钟机制还可以通过代表细胞生物能状态的代谢物和酶的时间丰度或活性进行微调,以使核心时钟蛋白被代谢物水平修饰,从而影响时钟驱动的基因表达。例如,负责激活时钟蛋白磷酸化的几种激酶,例如AMP激活蛋白激酶(AMPK)、核糖体蛋白S6激酶-β1(S6K1)和糖原合酶激酶3β(GSK3β)均受生物能和激素状态影响,因此强调代谢方面对生物钟功能的影响,细胞代谢状态是生物钟的关键(图2)。
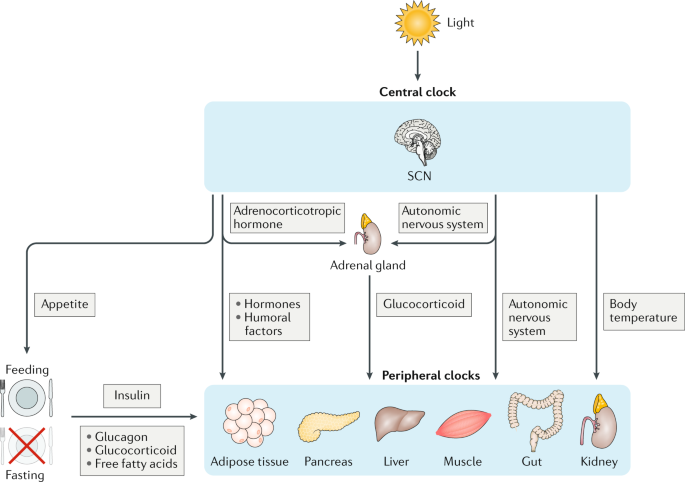
图一 生物钟网络
肿瘤自主时钟破坏
与正常组织相比,肿瘤细胞的昼夜节律似乎有些紊乱。转录组数据库荟萃分析显示,时钟基因在多种人类癌症中出现广泛的误表达。重要的是,时钟基因表达的错误程度似乎与各种癌症的分期或侵袭程度密切相关。如生物钟在霍奇金淋巴瘤晚期阶段被打乱。与其他所有乳腺癌相比,三组阴性乳腺癌的时钟基因误表达更多。而且时钟和细胞分化紧密交织,核心时钟在未分化的多能干细胞中未明显表现出来,但随着细胞分化和生物发育而逐渐表现出来。因此可以做出假设,一些生物钟调节因子可以发挥肿瘤抑制因子的作用。(图3)

图2 生物钟的分子机制
CLOCK和BMAL1
BMAL1低表达与胰腺癌、黑色素瘤和结直肠癌的进展、低分化和不良预后相关。过表达BMAL1能够减弱人类肝癌、骨肉瘤、白血病、结直肠癌和卵巢癌的肿瘤生长,而BMAL1的沉默减少了细胞凋亡、增强了细胞增殖和人骨肉瘤和小鼠结肠癌细胞肿瘤生长。同样,CLOCK蛋白水平在肾母细胞肿瘤中也表达减少,但CLOCK基因在肾母细胞瘤中的表达与在正常肾组织中表达相似,这表明CLOCK蛋白的转录后调控可依据其转录活性而降低。值得注意的是,肾母细胞瘤是由正常肾细胞去分化引起的,这说明肿瘤的发生或去分化与正常生物钟破坏有关。有趣的是,CLOCK的特定单核苷酸多态性(SNP)也与乳腺癌风险增加相关。此外,结直肠癌中也检测到CLOCK基因的突变,并且野生型CLOCK水平的重建可防止紫外线引起的DNA损伤。这都表明CLOCK基因组的突变可能与肿瘤发生相关。
PER家庭
PER蛋白构成生物钟的昼夜节律输出端,并充当CLOCK–BMAL1的阻遏物。与CLOCK–BMAL1通常在肿瘤中活性降低相一致,编码PER家族蛋白的基因在很多肿瘤中被抑制。PER1在人神经胶质瘤、胃癌、非小细胞肺癌、乳腺癌、前列腺癌和结肠癌中被抑制;PER2在脑、肺、胃、乳腺癌、淋巴瘤和白血病中被抑制,并与大肠癌的肝转移有关;PER3在结肠直肠癌中被抑制。PER2和PER3的表达增加与抑制肿瘤生长、增加肿瘤的分化或凋亡、减少淋巴结转移和延长无转移生存期有关。目前研究认为,昼夜节律在癌症发展过程中被抑制,并且这种抑制在功能上是相关的。

图3:肿瘤组织的生物钟
PER似乎也通过与非时钟转录因子相互作用而发挥抗肿瘤作用。外源表达PER1有可能成为新的治疗策略。PER1与雄激素受体结合并抑制雄激素受体介导的基因激活和生长,PER2促进雌激素受体降解,而沉默PER2能稳定乳腺癌细胞系中的雌激素受体,该受体参与乳腺癌的病理生理。PER2还可以通过与TF OCT1(也称为POU2F1)相互作用,招募抑制性复合物来充当转录共抑制子,从而抑制上皮细胞的生长。此外,PER2能够与人肠癌细胞中的肿瘤抑制物p53相互作用,促进p53易位进入细胞核并随后抑制肿瘤发生。因此,PER似乎执行了许多独立于时钟的功能,从而促进了抗肿瘤发生程序。作为Per1和Per2基因可以被非时钟的TF(如GR和CREB)诱导,这提示糖皮质激素可以通过特异性上调PER编码基因帮助抑制癌症进展的。
CRY家族
肿瘤细胞中CRY蛋白的行为似乎比PER更复杂。Cry1的表达主要受ROR–REV-ERB轴的控制。肠癌细胞中CRY1表达的增加与肿瘤进展和预后不良有关,大概反映了CLOCK-BMAL1活性的降低。Cry 2由CLOCK–BMAL1直接调节。CRY2的表达在乳头状和滤泡性甲状腺癌中受到抑制,而CRY2的高表达与乳腺癌无转移和更长生存期相关。重要的是,同时敲除两个CRY基因会导致CLOCK–BMAL1的激活,从而防止癌症发展并改善p53突变小鼠的预后。这表明,只要在昼夜节律周期中高度激活CLOCK-BMAL1复合物,节律紊乱对肿瘤发生的负面影响就可能很小。需要进一步的研究来识别CLOCK–BMAL1的昼夜节律对肿瘤发生的具体贡献。和PER相似,CRYS也可以通过靶向非时钟的TF具有非时钟依赖性功能。
生物钟与新陈代谢
如前所述,癌细胞中的正常生物钟似乎经常受到干扰,从而促进肿瘤的增殖代谢。因此,宿主生物体中昼夜节律的长期破坏有可能促进肿瘤的发生发展。昼夜周期深刻影响了下丘脑上眼睑上核(SCN)的中央时钟,而进食禁食节律则带动了周围组织的昼夜节律。SCN衍生的日常信号支配着松果体,肾上腺,脾脏和骨髓等外周组织的每日生理水平,其中褪黑激素,糖皮质激素,淋巴细胞计数和造血功能的昼夜水平受到严格调节。据推测,通过这些因素以及其他神经和体液网络,由于慢性光周期改变或饮食习惯不规律造成的昼夜节律紊乱可能导致潜在的致瘤性时钟失调。事实上,轮班工作或夜间饮食等生活方式导致的昼夜颠倒是患各种癌症的危险因素,包括乳腺癌、子宫内膜癌、前列腺癌、肺癌、结肠癌、肝癌症和非霍奇金淋巴瘤。特别是与性类固醇激素相关癌症的相关性是有据可查的,这与上述PER在雄激素或雌激素信号传导相关。然而,昼夜节律与某些类型的癌症(例如结肠癌)之间的联系可能是由于继发性作用所致。例如,昼夜节律紊乱可促进肥胖,肥胖本身就是癌症的危险因素。研究已经阐明了多种潜在机制通过昼夜节律破坏促进肿瘤生长(图4)。

图4 宿主生物的生物节律与癌症自主生物钟之间的交流
褪黑激素
SCN中的中央昼夜节律起搏器起着肿瘤抑制器的基本作用。例如,小鼠去除SCN后,其昼夜活动节律被打乱,这促进了骨肉瘤和胰腺腺癌的生长。SCN去除后诱导的这种致癌作用可以通过改变曝光时间得以实现。已经观察到长期倒时差或持续光照足以促进小鼠中骨肉瘤和乳腺癌及大鼠中肝癌的生长。通过夜间使用昏暗的光线来破坏昼夜节律,会损害宿主的新陈代谢,诱发例如高血糖和高胰岛素血症,从而导致大鼠的需氧糖酵解增强和肿瘤增殖。持续的光照和黑暗通过抑制褪黑激素的合成促进肝癌的生长和乳腺上皮细胞中DNA合成。褪黑激素是松果体分泌的一种完整的昼夜节律激素,它增加了线粒体的氧化并阻碍了肿瘤脂质的代谢和生长。褪黑激素水平在SCN中央起搏器的控制下,通过与多突触途径的连接而受到光的抑制。流行病学研究证实了褪黑激素可能具有的肿瘤抑制特性,褪黑激素水平与癌症风险成反比关系。给予褪黑激素能够减少乳腺雌激素水平和DNA合成,或通过模拟生理光照周期抑制肿瘤生长。此外,大鼠松果体功能的丧失也促进了乳腺肿瘤的生长,通过给以褪黑激素可以部分挽救这种效应。
糖皮质激素
糖皮质激素水平的昼夜节律被破坏也可能促进肿瘤发生。糖皮质激素是外周时钟的强大同步器。GR与编码PER家庭成员的基因启动子糖皮质激素应答元件(GRE)结合并诱导PER表达。虽然下丘脑-垂体-肾上腺轴主要支配肾上腺糖皮质激素生产,交感神经和肾上腺固有时钟也调节糖皮质激素水平。SCN去除破坏了糖皮质激素水平的昼夜节律,这可能影响时钟周期,并因此促进肿瘤发生。有趣的是,由于光照时间的长期变化而引起的小鼠乳腺肿瘤生长是在体温和糖皮质激素皮质激素水平异常循环之前,这说明全身性因素参与了肿瘤发生。鉴于GR被CLOCK介导的乙酰化作用所抑制,并和CRYS相互作用,可通过破坏CLOCK乙酰转移酶活性和CRY1、CRY2水平导致糖皮质激素生理节律紊乱。流行病学研究表明,糖皮质激素也可能导致与压力相关的癌症进展。糖皮质激素水平模式的改变可能会影响淋巴细胞功能,从而影响抗癌免疫力。
免疫功能
抵抗肿瘤细胞的免疫力是预防肿瘤发生的重要组成部分。血液中循环的淋巴细胞数量显示出昼夜节律性,去除SCN破坏了淋巴细胞计数的昼夜节律,这可能有助于癌细胞增殖。光照周期的频繁变化可导致大鼠免疫功能降低,并促进小鼠移植瘤生长,降低小鼠存活率。此外,长期倒时差引起的昼夜节律紊乱损害了抗癌自然杀伤细胞的溶细胞能力,导致大鼠肺肿瘤生长。
值得注意的是,源自SCN中央起搏器的周期性交感神经输入似乎对周围组织具有直接的肿瘤抑制作用,而与调节免疫功能无关。交感神经支配已被证明在多种类型的癌症进展、侵袭和转移中发挥重要作用。
肿瘤衍生因子与癌症化疗
除了昼夜节律的变化会影响宿主代谢和肿瘤的发展,荷瘤状态也可能从根本上干扰或重编程宿主新陈代谢的昼夜节律。事实上,癌症患者经常表现出行为和/或褪黑激素、皮质醇血浆水平和淋巴细胞计数昼夜节律紊乱。维持皮质醇或生活行为的昼夜正常节律与患者乳腺癌或结肠癌预后较好和生活质量相关。另外,研究发现患肺腺癌、乳腺癌和肠癌的小鼠可以远程设置其肝脏内的生物钟基因表达。这种暂时的器官间通信可以通过各种潜在的机制来解释。肿瘤经常使用有氧糖酵解,这会在血液中产生大量的乳酸。虽然肿瘤细胞可以使用自产乳酸盐作为三羧酸循环的燃料源,循环乳酸也可由肝脏还原代谢为葡萄糖,但为了代谢更多的乳酸,肝脏需要改变昼夜节律基因组程序,例如,患肺肿瘤的小鼠的肝脏会表现出独有的非常丰富的生物钟基因亚群。此外,肿瘤通常分泌细胞因子,例如白细胞介素6(IL-6),这可能会调节远端器官的昼夜节律代谢。例如,IL-6影响携带肺肿瘤的小鼠肝脏中的胰岛素信号传导途径。乳腺癌可以通过小鼠的交感神经引起降钙素/食欲肽神经元异常活动,从而导致睡眠-觉醒周期的破坏和葡萄糖代谢的改变。尤为重要的是,睡眠-觉醒周期可以与昼夜节律独立开来。因此,即使在明暗周期下,睡眠剥夺也破坏了大脑中突触蛋白表达的日常节律。睡眠片段化似乎与肿瘤发展相关联,昼夜破坏和睡眠障碍至少部分独立地和协同地促进肿瘤发生。
此外,抗癌化学疗法也可能有助于破坏宿主的昼夜节律代谢。外源性、长效糖皮质激素通常作为化疗方案的一部分,这可引起每日葡萄糖稳态的失调。与化学疗法有关的心理压力以及诸如恶心和呕吐等不良反应可能会影响睡眠和宿主的昼夜节律。这些观察结果表明,肿瘤向宿主生物的生理和代谢传递了深远的信号。总的来说,可以推断出肿瘤或由治疗引起的宿主代谢的昼夜节律紊乱进一步支持了癌症的进展,从而在肿瘤自主钟和宿主代谢的昼夜节律之间产生了恶性循环。
恢复正常代谢生物钟可作为潜在的癌症治疗方法
研究证明,纠正患瘤小鼠紊乱的代谢生物钟,可以恢复宿主生物体的代谢节奏,抑制肿瘤生长。同样,与随意喂养相比,定时摄入食物可抑制小鼠胰腺腺癌的生长。在该模型中,肿瘤生长的抑制伴随着原生物钟紊乱的癌细胞中生物钟基因的恢复,这表明宿主代谢的变化可能会驱动肿瘤细胞的昼夜节律,从而导致肿瘤抑制。时间限制性喂食的有益效果可能部分地归因于禁食相,空腹有利于正常生物钟的维持和防止肿瘤发生。该概念可以进一步扩展到其他代谢途径。老年小鼠的热量限制能够诱导生物钟基因高表达。同样,在高脂饮食诱导的NAD +耗竭的肝脏中使用其前体代谢产物烟酰胺单核苷酸(NMN)补充NAD +可以恢复核心时钟基因的表达。尽管需要进一步的研究,以解决衰老或饮食引起的肥胖症中昼夜节律的减弱与肿瘤发生有关的程度,但这些针对昼夜节律的代谢干预措施代表了减慢肿瘤生长的替代策略。总而言之,这些发现表明,以宿主和癌细胞之间的昼夜代谢节律为靶向的治疗方法可能为癌症治疗开辟新的途径。
结论和未来方向
昼夜节律的紊乱导致生物钟功能的破坏,并且所产生的利肿瘤性新陈代谢可能增加肿瘤发生发展的可能性。尽管我们对生物钟系统和肿瘤发生有基本的了解,但仍然存在许多悬而未决的问题。例如,已知在各种癌症中染色质会发生变化,但实际上昼夜节律相关转录因子(例如CLOCK–BMAL1)在基因上的基因组占有率是否在癌症中也发生了变化,这一点尚待探索。如果是这样,就必须仔细分析上皮-间质肿瘤在转化或转移过程中,这些变化是否与昼夜节律的相关基因和蛋白质表达在功能上相关。尚不清楚是否存在非生物钟依赖的特定染色质状态。另外,仍然缺乏关于不同类型癌症中细胞自主生物钟表达和功能的独特模式的证据。此外,缺少关于癌症和多能干细胞之间是否存在共同的核心时钟信号的信息,这些信号可能在细胞分化的某个阶段发生分化。所有这些问题对于未来治疗策略的发展尤其重要,其中可能包括恢复功能性时钟和逆转癌症表观基因组的可塑性。
“论肿道麻”点评
视交叉上核(SCN,suprachiasmaticnucleus)是哺乳动物包括人最重要的生物钟,调节生物体内生化、生理和行为功能的内在周期震荡,SCN通过神经元连接以及机体分泌的可扩散分子,向神经内分泌系统和植物神经系统中的其他丘脑中心发出昼夜节律信号。哺乳动物50-80%的蛋白质编码基因由生物钟控制,其中包括一些对细胞分裂时序至关重要的基因。长期昼夜节律失调影响细胞周期、细胞程序性死亡以及DNA损伤,促进肿瘤细胞逃逸,提高肿瘤增殖速率。
此综述着重阐述了分化与未分化细胞中生物钟基因和蛋白质的表达模式;决定增殖代谢的核心时钟和非时钟转录因子(TF);以及肿瘤与宿主之间的相互影响。生物钟可与非时钟转录因子协同引导肿瘤细胞的代谢过程,以控制营养物质和代谢物如何被利用,从而最终影响细胞分化和增殖。强调细胞代谢影响生物钟功能,肿瘤细胞的昼夜节律紊乱,而生物钟调节因子可以发挥肿瘤抑制因子的作用。上调PER编码基因、激活CLOCK-BMAL1复合物可降低节律紊乱对肿瘤的负面影响。昼夜节律紊乱影响褪黑激素,糖皮质激素,淋巴细胞计数和造血功能的水平,促进肿瘤细胞生长;患者荷瘤状态和抗肿瘤化学治疗引起的宿主代谢的昼夜节律紊乱进一步支持了癌症的进展,从而形成恶性循环。因此,以宿主和癌细胞之间的昼夜代谢节律为靶向的治疗方法可能为癌症治疗开辟新的途径;了解生物钟机制与化疗药物治疗靶点之间的特定相互作用优化肿瘤防治策略;利用昼夜节律和生物钟来预防和治疗肿瘤有望成为新的研究方向。
编译 潘波,述评 陈蔚
审校:张军,缪长虹
原始文献:Kinouchi K, Sassone-Corsi P. Metabolic rivalry: circadian homeostasis and tumorigenesis. Nat Rev Cancer. 2020;20(11):645-661. doi:10.1038/s41568-020-0291-9

不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










