


















西农 | SBB:作物覆盖和施肥处理下苹果园不同粒径土壤团聚体中丰富、稀有细菌和真菌亚群落的组装




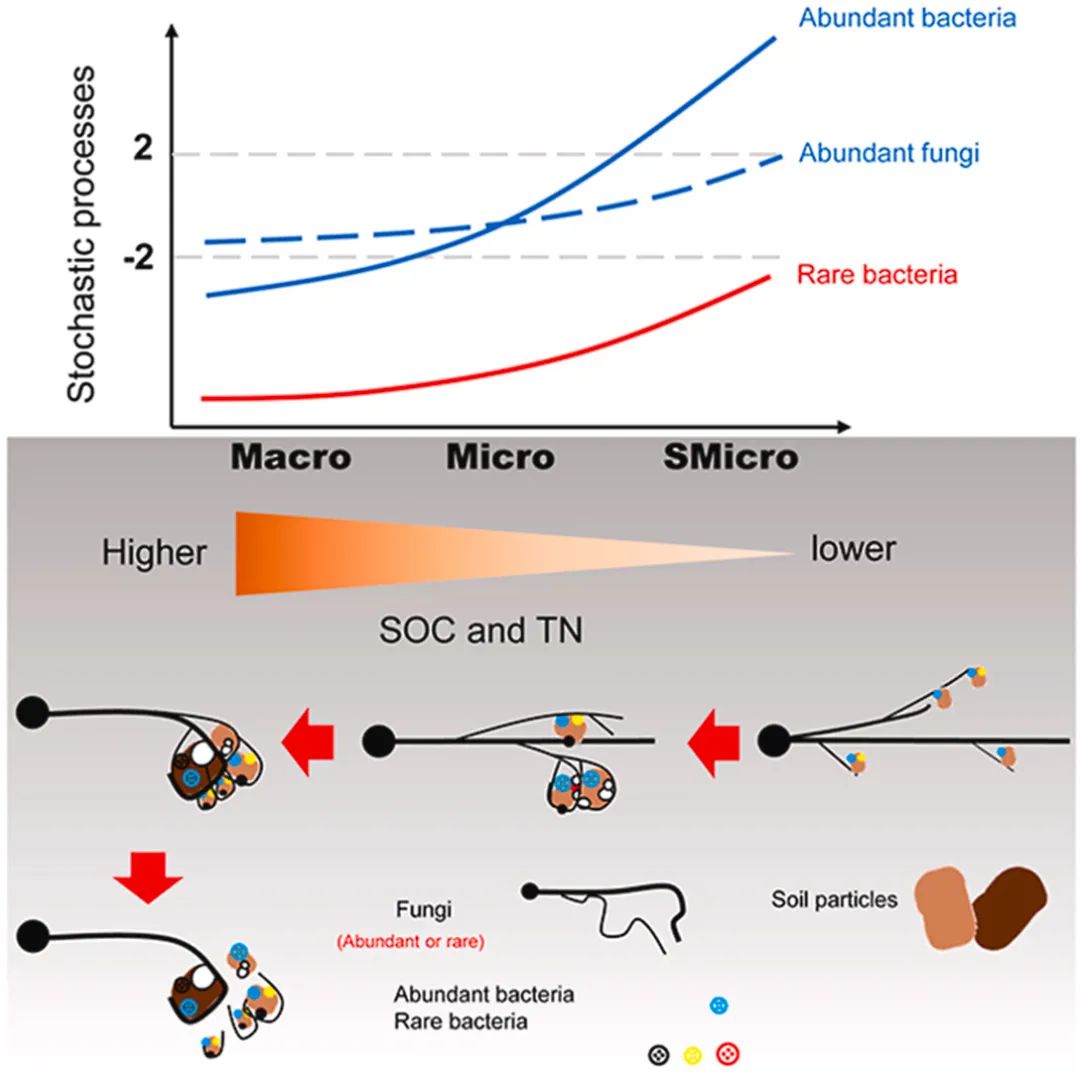
丰富真菌群落的组装以随机过程为主,而不同粒径团聚体中稀有真菌群落的组装以同质选择和有限扩散为主导。
编译:微科盟清韵,编辑:微科盟木木夕、江舜尧。
微科盟原创微文,欢迎转发转载。
土壤细菌和真菌群落通常由“丰富生物圈”和“稀有生物圈”组成,它们共存于土壤团聚体中,形成一个复杂的种间相互作用系统。然而,由于团聚体的空间隔离,丰富和稀有细菌和真菌群落所表现出的不同的组装过程仍然不清楚。本研究量化了覆盖作物和施肥处理的苹果园中不同粒径大小的土壤团聚体中与丰富和稀有的细菌和真菌亚群落组装相关的随机性和确定性过程。在丰富细菌群落的组装中,变量选择的重要性从大团聚体增加到小型微团聚体。随着土壤有机碳和总氮含量的降低,稀有细菌群落的组装过程经历了从同质选择到弱同质选择的转变。土壤团聚体中丰富和稀有细菌群落在扩散上都不受限制。丰富真菌群落的组装以随机过程为主,而不同粒径团聚体中稀有真菌群落的组装以同质选择和有限扩散为主导。丰富和稀有细菌和真菌群落的组成也受到土壤团聚体和农业措施的影响。与稀有细菌和真菌相比,细菌和真菌中含量最高的前20个OTUs大多与大团聚体数量有关,而丰富真菌的OTUs中也有部分与微团聚体和小型微团聚体有关。本研究为今后的实验验证提供了先验假设,从而为理解不同粒径土壤团聚体中真菌和细菌群落组成变化的机制提供了系统的方法。
论文ID
原名:Assembly of abundant and rare bacterial and fungal sub-communities indifferent soil aggregate sizes in an apple orchard treated with cover crop and fertilizer
译名:作物覆盖和施肥处理下苹果园不同粒径土壤团聚体中丰富、稀有细菌和真菌亚群落的组装
期刊:Soil Biology and Biochemistry
IF:5.795
发表时间:2021.03
通讯作者:翟丙年
通讯作者单位:西北农林科技大学
实验设计和数据分析
结果







讨论
结论


不感兴趣
看过了
取消

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










