


















一阴一阳之谓道:顾伟实验室总结p53基因在肿瘤代谢中的复杂作用




p53 是一个拥有广泛而强大功能的抑癌基因。






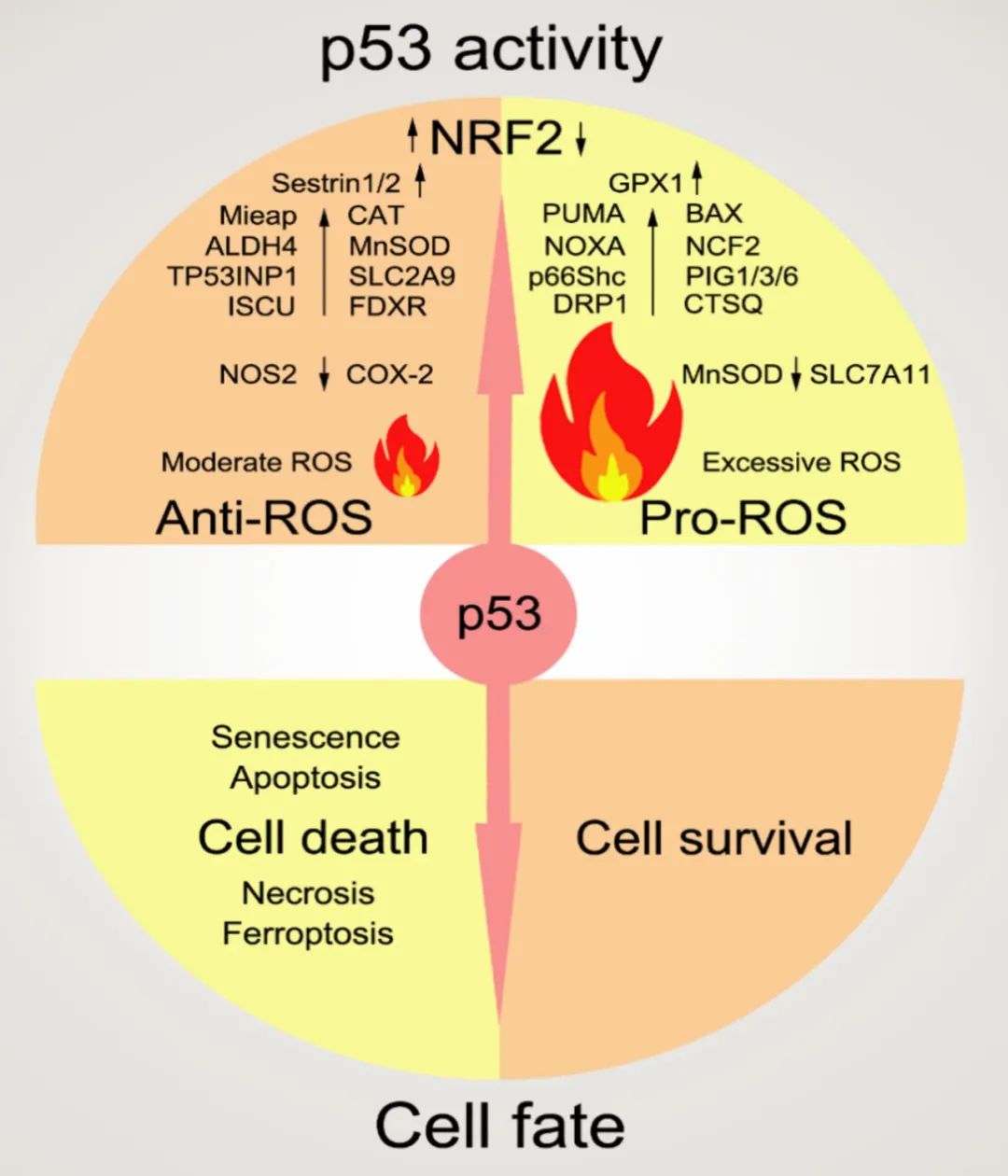
需要指出的是,PFK1是糖酵解过程中的限速酶。p53可以通过激活靶基因TIGAR的表达来抑制PFK1活性,进而下调糖酵解的速率。但是抑制PFK1会导致PPP通路的激活,反而有利于癌细胞的存活。而在某些特殊情况下,比如骨骼肌和心肌细胞里,p53却是激活PGM来促进糖酵解的。 (p53的代谢调节功能非常复杂,很多时候其效果是context-dependent的,常常会有矛盾的现象出现,这将在下文中多次出现。文末会对该现象进行总结讨论。)







不感兴趣
看过了
取消
本文由“健康号”用户上传、授权发布,以上内容(含文字、图片、视频)不代表健康界立场。“健康号”系信息发布平台,仅提供信息存储服务,如有转载、侵权等任何问题,请联系健康界(jkh@hmkx.cn)处理。
关键词:
p53,肿瘤,基因,代谢,细胞,功能,调节

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










