


















综述 | Water Research:水生系统中细菌的共凝聚




多种微生物群落在多个研究领域中的重新崛起和多学科研究方法的最新进展为人们理解共凝聚在固定群落的建立和行为中的作用提供了更多的兴趣。
论文ID
原名:Bacterial coaggregation in aquatic systems
译名:水生系统中细菌的共凝聚
期刊:Water Research
IF:9.130
发表时间:2021.3.10
通讯作者:Manuel Simões
通讯作者单位:葡萄牙波尔图大学化学工程系学工程系工程系
综述框架

主要内容
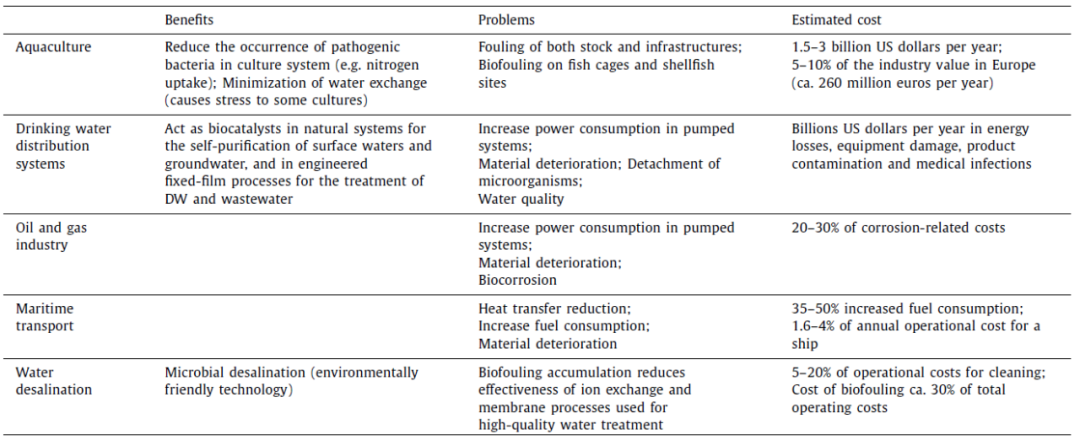





结论

不感兴趣
看过了
取消
本文由“健康号”用户上传、授权发布,以上内容(含文字、图片、视频)不代表健康界立场。“健康号”系信息发布平台,仅提供信息存储服务,如有转载、侵权等任何问题,请联系健康界(jkh@hmkx.cn)处理。
关键词:
生物膜,微生物,细菌,菌株

人点赞

人收藏

打赏
不感兴趣
看过了
取消













 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










