


















文献精读 | 敲除施万细胞中的钙调神经磷酸酶对髓鞘化无影响,但可降低周围损伤后自噬并延缓鞘磷脂形成





前言
在周围神经系统(PNS)中,Schwann细胞(SCs)将轴突包裹在同心的脂质膜双层中形成髓磷脂。髓磷脂充当电信号沿轴突传播的绝缘层。周围神经的发育取决于SC与轴突之间的信号传导。早期的SC,称为SC前体(SCP)为轴突提供营养支持,而SCP则需要轴突发出NRG1使其分化为SC,并随后产生髓磷脂。
NRG1结合SC上的ErbB2 / 3受体,并激活下游的髓鞘再生途径,包括Ras /细胞外信号调节激酶1/2(ERK1 / 2)和磷脂酰肌醇-3-激酶(PI3K)/ AKT途径。这些途径可在体内和体外改变髓鞘厚度。先前的文献表明,NRG1还可以激活钙调神经磷酸酶。钙调磷酸酶是一种被钙激活的丝氨酸/苏氨酸蛋白磷酸酶,由钙调磷酸酶A和钙调磷酸酶B两个亚基组成,并经常被研究其在NFAT转录因子(活化T细胞的核因子)的去磷酸化和激活中的作用。
体内神经细胞中调节亚基CnB的缺失会抑制钙调磷酸酶活性,并在胚胎和围产期引起径向分选延迟和髓鞘减少。为了评估钙调神经磷酸酶仅在SC中的作用,研究者使用CnB-floxed和P0-Cre小鼠(CnBscko)生成了SC条件性CnB基因敲除小鼠,评估出生后的外周髓鞘形成。出人意料的是,CnBscko小鼠的髓鞘缺损最少,髓鞘蛋白表达没有减少,提示钙调神经磷酸酶在发育性髓鞘形成中的作用并非关键。尽管对发育中的髓鞘形成几乎没有影响,但钙调神经磷酸酶在损伤后的SC中具有作用。
与中枢神经系统(CNS)不同,PNS在受伤后具有惊人的再生能力。在受到伤害的几分钟内,轴突开始在损伤部位远端退化,这一过程称为Walleria退化。这一步是由轴突引发的,并触发了SC的转分化过程,从前髓鞘的SC转变为“脱髓鞘”的SC,然后是“修复”的SC。SC首先通过肌动蛋白聚合的变化使它们的髓鞘破裂。然后,髓磷脂的这种碎裂作为髓磷脂卵形夹入细胞质中。
SCs可通过多种平行途径分解并消化自身的髓磷脂,从而使轴突神经突再生。研究表明SCs可以通过自噬,受体介导的吞噬以及通过MLKL破坏髓鞘,清除碎片。从遗传学或药理学角度抑制SC的自噬,会减慢髓鞘蛋白的清除速度,使残留的髓鞘数量增加。通常,抑制mTOR将激活自噬。然而,损伤后mTOR活性在SC中上调,这表明髓吞噬细胞的激活是通过非mTOR途径发生的。
钙调神经磷酸酶为自噬的新型调节剂。与此同时,消除CnB可以破坏损伤后的髓磷脂清除能力;缺乏CnB的SCs,自噬功能降低。
总之,本研究表明钙调神经磷酸酶是损伤后自噬的新型调节剂,而不是以前认为的重要的髓鞘生成因子。
01
特异性消除SCs中的CnB引起轻微的径向分选延迟,但无髓鞘减少
从条件性敲除CnB的P5-P6幼鼠的坐骨神经的雪旺细胞中CnB减少(图1a)的同时,钙调神经磷酸酶的酶亚单位钙调神经磷酸酶A(CnA)也减少(图1a),表明CnB在CnA亚基的稳定性中发挥了作用。
接下来,我们分析了髓鞘形成(P1),活动性髓鞘形成(P10),髓鞘形成后(P28)和成年期(6 mo)时,小鼠的坐骨神经形态。在所有这些时间点,髓鞘厚度和髓磷脂蛋白表达均无差异(图1,d,e,c)。但是,在CnBscko小鼠中,轴突径向分选过程存在一定程度的延迟(图1f,g)。与WT相比,P28和6个月时无髓鞘轴突的数量增加了(图1h)。有趣的是,在6个月大的CnBscko小鼠中,物髓鞘纤维的数量大于P28 CnBscko小鼠中的数量,这表明其中一些纤维已经脱髓鞘而不是从未被髓鞘化(图1h)。这些数据表明钙调神经磷酸酶的损失会引起轻度的径向分选延迟,但总的钙调神经磷酸酶在发育性髓鞘形成和实现正确的髓磷脂厚度方面的作用有限。

图1
02
CnB-scko小鼠坐骨神经中SC细胞的增殖和分化不受影响
通过免疫荧光染色评估P10 CnB-scko小鼠中KROX20的表达。与WT相比,我们没有观察到核表达的降低(图2a,b)。追溯发育过程中,CnBscko小鼠神经中的KROX20蛋白表达亦没有降低(图2d,e)。CnB-scko坐骨神经中的前髓鞘转录因子OCT6也没有变化(图2a,c)。综上敲除SC前体细胞中的CnB的不会影响其成熟和髓磷脂转录因子KROX20的上调。由于钙调神经磷酸酶被认为是NRG1的下游,我们分析了Schwann细胞中NRG1下游的其他已知途径,以确定是否存在补偿钙调神经磷酸酶缺失的机制。P1,P10,P28和6月龄坐骨神经中ERK的磷酸化(图2f)和P1,P10和P28坐骨神经中的AKT的磷酸化的结果显示WT和CnB-scko在P1,P10,P28和6 mo之间没有差异(图2f,g,h,i)。以上提示并不存在补偿机制。量化P3 WT和CnB-scko坐骨神经中Ki67阳性SC细胞(SOX10)的百分比发现在增值方面不存在差异(图2j-1)。

图2
03
SC细胞中CnB的丢失会延迟损伤后的细胞变性
受伤部位远端的神经会发生Wallerian变性。这包括一系列变化,包括远端变性和碎片清除后轴突和髓鞘的再生。受伤后(dpc)第7天,在CnB-scko坐骨神经中未受伤部分和变性轴突周围都包裹着更多的髓鞘(图3a-b)。有趣的是, 3dpc的CnB-scko小鼠中的轴突变性更少(图3c,d)。这些数据表明髓鞘清除率和轴突变性之间可能存在联系。
qPCR结果显示趋化剂MCP-1在3dpc时的表达没有差异,表明钙调神经磷酸酶的丧失不影响SC细胞和成纤维细胞在损伤后募集巨噬细胞的能力(图3e)。此外,受伤神经远端F480 +细胞数量在3dpc和7dpc时间点均无差异(图3f,g)。
建立坐骨神经变性的离体模型来评估损伤后CnB丢失的Schwann细胞的自主效应(图4a)。与WT神经相比, 培养12天后,CnB-scko坐骨神经的髓鞘和轴突更完整(图4b)。同样,原代SC保留在培养基中以消化自身的髓磷脂,并在1、4、7天(div)进行分析(图4c)。培养7天后,CnB-scko幼鼠中分离的原代SC中含有 P0髓磷脂蛋白的比例更高(图4d)。综上,在CnB-scko小鼠在7dpc时所见的髓鞘清除率受损不是由于免疫细胞浸润的差异,而是由于SC氏细胞自我缺陷。

图3

图4
04
钙调神经磷酸酶不调节损伤后SC细胞的分化和增殖
在受伤后的几个小时内,SC细胞增殖并开始转分化为修复的SC。分化会阻止SC细胞降解髓磷脂、支持轴突再生和髓鞘化新轴突的能力,从而阻止受伤后的恢复。分化的主调节因子c-JUN在CnBscko远端残端中的表达水平不变。(图5a-d,e,5g)。mTORC1位于 c-JUN的上游,在3和7 dpc时,CnBscko和WT远端神经之间的mTOR活性没有显着差异(图5a-d)。此外, OCn6(标记未成熟SC)和P-ERK表达(免疫细胞募集,再生)均在CnB-scko小鼠的神经远端残端中被激活。最后, SC细胞的增殖在CnB-scko小鼠也未发生变化(图5f,5h)。总之,这些数据表明损伤后,SC在CnB-scko神经中正确转分化;在CnB-scko SC中观察到的表型是由于髓鞘清除的问题,而不是由于SC细胞转变为修复表型。
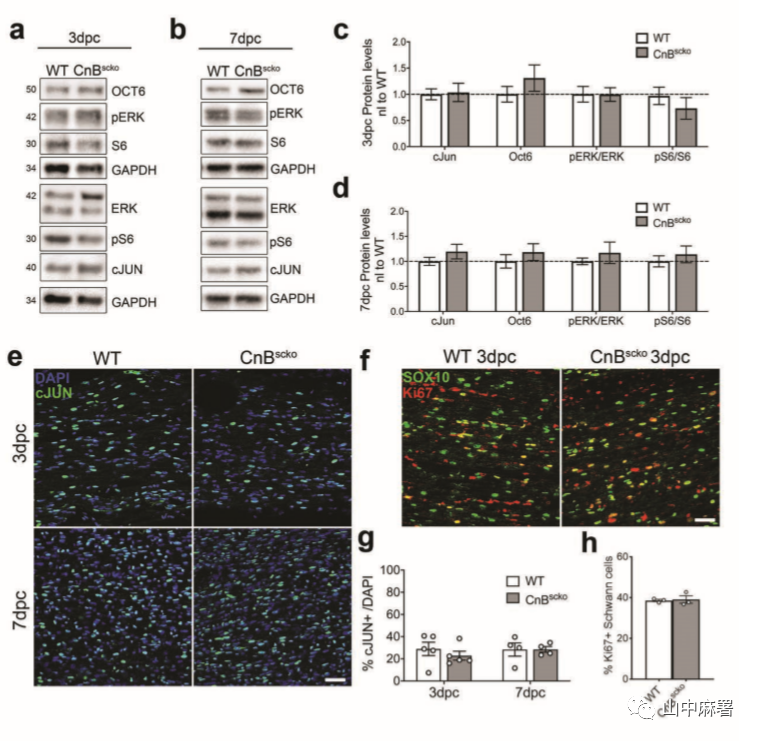
图5
05
CnB的缺失会钝化TFEB激活并损害自噬作用
SC细胞在损伤后使用自噬作用来清除髓磷脂碎片。条件性敲除SC中Atg7(自噬相关基因7)可防止自噬,其表型类似于CnBscko神经损伤。这种相似性得以延伸:在ATG7scko神经中,SC的转分化和增殖是正常的。因此,假设钙调神经磷酸酶,一种自噬的新型调节剂,可能参与了损伤后SC中的髓鞘吞噬。对SC的自噬通量进行测量,用氯化铵(NH4Cl)处理分离的Schwan细胞,并测量LC3II表达。氯化铵通过抑制自噬体和溶酶体的融合,防止其降解并引起自噬体的蓄积。通过测量LC3II蛋白(自噬小体的一个组成部分)来进行量化。结果显示在WT的SC中,用NH4Cl处理3小时会导致LC3II表达的显著增加,而在CnBsckoSC中未观察到。此外,未经处理的CnBsckoSC已经表达了高水平的LC3II(图6a)。经处理后,CnBscko细胞中LC3II的表达没有增加,这表明CnBscko SC在这段时间内没有产生更多的自噬体。缺少CnB的SC出现自噬通量下降(图6b)。综上所述, CnB的丢失会阻止SC在损伤后的自噬活性,并损害髓磷脂的清除。
钙调神经磷酸酶是通过调控转录因子EB(TFEB)来调控溶酶体生物发生和自噬的调节然而,关于TFEB在神经损伤中的作用知之甚少。因此,为了确定神经损伤后TFEB是否被激活,首先分析了WT损伤神经近端和远端中TFEB的表达(图6c)。在第7dpc时,远端的核TFEB明显增加(图6d)。随后在3dpc和7dpc CnBscko小鼠的远端神经中重复了该实验(图6e)。与3dpc的WT相比,CnBscko神经中TFEB+ SC的数量下降,而TFEB+巨噬细胞数量不变(图6f)。TFEB mRNA的结果于此一致,并与TFEB调节自身转录的事实相符(图6g)。相反, 7dpc时间点各组之间不再存在差异(图6h)。这可能与7dpc是TFEB的重建是其他激活TFEB的磷酸酶的补偿作用。总之,自噬的缺陷可能是CnBscko神经中TFEB激活被抑制的结果。

图6
06
SC中的钙调神经磷酸酶对于受伤后的再髓鞘化不是必需的
为确定CnBscko神经中观察到的TFEB和自噬激活的延迟是否影响轴突再生和再髓鞘化,对 21dpc时间点的WT和CnBscko神经(图7a)。尽管髓鞘清除延迟,但在21dpc时,CnBscko坐骨神经与WT神经相当:髓鞘厚度(图7b),髓鞘纤维数量,无髓鞘纤维(蓝色箭头)或具有髓鞘碎片的细胞(红色箭头)的数量没有差异(图7c)。这些数据表明钙调神经磷酸酶在损伤后髓磷脂的变性和清除中起作用,而不是在轴突再生和髓鞘再生中起作用。综上所述,可以假设存在其他未知的SC自主机制,这些机制在损伤后的髓磷脂消化中起作用,并可以补偿这些模型中自噬的减少,从而实现适当的髓鞘再生。

图7
结论
钙调神经磷酸酶对于髓鞘形成非必需,但参与损伤后髓鞘清除过程。
原始文献:
Reed CB, Frick LR, Weaver A, et al. Deletion of Calcineurin in Schwann Cells Does Not Affect Developmental Myelination, But Reduces Autophagy and Delays Myelin Clearance after Peripheral Nerve Injury. J Neurosci. 2020;40(32):6165-6176. doi:10.1523/JNEUROSCI.0951-20.2020
本文由“山中麻薯”授权转载


人点赞

人收藏

打赏









 010-82736610
010-82736610
 股票代码: 872612
股票代码: 872612
 京公网安备 11010802020745号
京公网安备 11010802020745号










